- Patient Care & Health Information
- Diseases & Conditions
- Coronavirus disease 2019 (COVID-19)
COVID-19, also called coronavirus disease 2019, is an illness caused by a virus. The virus is called severe acute respiratory syndrome coronavirus 2, or more commonly, SARS-CoV-2. It started spreading at the end of 2019 and became a pandemic disease in 2020.

- Coronavirus
Coronaviruses are a family of viruses. These viruses cause illnesses such as the common cold, severe acute respiratory syndrome (SARS), Middle East respiratory syndrome (MERS) and coronavirus disease 2019 (COVID-19).
The virus that causes COVID-19 spreads most commonly through the air in tiny droplets of fluid between people in close contact. Many people with COVID-19 have no symptoms or mild illness. But for older adults and people with certain medical conditions, COVID-19 can lead to the need for care in the hospital or death.
Staying up to date on your COVID-19 vaccine helps prevent serious illness, the need for hospital care due to COVID-19 and death from COVID-19 . Other ways that may help prevent the spread of this coronavirus includes good indoor air flow, physical distancing, wearing a mask in the right setting and good hygiene.
Medicine can limit the seriousness of the viral infection. Most people recover without long-term effects, but some people have symptoms that continue for months.
Typical COVID-19 symptoms often show up 2 to 14 days after contact with the virus.
Symptoms can include:
- Shortness of breath.
- Loss of taste or smell.
- Extreme tiredness, called fatigue.
- Digestive symptoms such as upset stomach, vomiting or loose stools, called diarrhea.
- Pain, such as headaches and body or muscle aches.
- Fever or chills.
- Cold-like symptoms such as congestion, runny nose or sore throat.
People may only have a few symptoms or none. People who have no symptoms but test positive for COVID-19 are called asymptomatic. For example, many children who test positive don't have symptoms of COVID-19 illness. People who go on to have symptoms are considered presymptomatic. Both groups can still spread COVID-19 to others.
Some people may have symptoms that get worse about 7 to 14 days after symptoms start.
Most people with COVID-19 have mild to moderate symptoms. But COVID-19 can cause serious medical complications and lead to death. Older adults or people who already have medical conditions are at greater risk of serious illness.
COVID-19 may be a mild, moderate, severe or critical illness.
- In broad terms, mild COVID-19 doesn't affect the ability of the lungs to get oxygen to the body.
- In moderate COVID-19 illness, the lungs also work properly but there are signs that the infection is deep in the lungs.
- Severe COVID-19 means that the lungs don't work correctly, and the person needs oxygen and other medical help in the hospital.
- Critical COVID-19 illness means the lung and breathing system, called the respiratory system, has failed and there is damage throughout the body.
Rarely, people who catch the coronavirus can develop a group of symptoms linked to inflamed organs or tissues. The illness is called multisystem inflammatory syndrome. When children have this illness, it is called multisystem inflammatory syndrome in children, shortened to MIS -C. In adults, the name is MIS -A.

When to see a doctor
Contact a healthcare professional if you test positive for COVID-19 . If you have symptoms and need to test for COVID-19 , or you've been exposed to someone with COVID-19 , a healthcare professional can help.
People who are at high risk of serious illness may get medicine to block the spread of the COVID-19 virus in the body. Or your healthcare team may plan regular checks to monitor your health.
Get emergency help right away for any of these symptoms:
- Can't catch your breath or have problems breathing.
- Skin, lips or nail beds that are pale, gray or blue.
- New confusion.
- Trouble staying awake or waking up.
- Chest pain or pressure that is constant.
This list doesn't include every emergency symptom. If you or a person you're taking care of has symptoms that worry you, get help. Let the healthcare team know about a positive test for COVID-19 or symptoms of the illness.
There is a problem with information submitted for this request. Review/update the information highlighted below and resubmit the form.
From Mayo Clinic to your inbox
Sign up for free and stay up to date on research advancements, health tips, current health topics, and expertise on managing health. Click here for an email preview.
Error Email field is required
Error Include a valid email address
To provide you with the most relevant and helpful information, and understand which information is beneficial, we may combine your email and website usage information with other information we have about you. If you are a Mayo Clinic patient, this could include protected health information. If we combine this information with your protected health information, we will treat all of that information as protected health information and will only use or disclose that information as set forth in our notice of privacy practices. You may opt-out of email communications at any time by clicking on the unsubscribe link in the e-mail.
Thank you for subscribing!
You'll soon start receiving the latest Mayo Clinic health information you requested in your inbox.
Sorry something went wrong with your subscription
Please, try again in a couple of minutes
COVID-19 is caused by infection with the severe acute respiratory syndrome coronavirus 2, also called SARS-CoV-2.
The coronavirus spreads mainly from person to person, even from someone who is infected but has no symptoms. When people with COVID-19 cough, sneeze, breathe, sing or talk, their breath may be infected with the COVID-19 virus.
The coronavirus carried by a person's breath can land directly on the face of a nearby person, after a sneeze or cough, for example. The droplets or particles the infected person breathes out could possibly be breathed in by other people if they are close together or in areas with low air flow. And a person may touch a surface that has respiratory droplets and then touch their face with hands that have the coronavirus on them.
It's possible to get COVID-19 more than once.
- Over time, the body's defense against the COVID-19 virus can fade.
- A person may be exposed to so much of the virus that it breaks through their immune defense.
- As a virus infects a group of people, the virus copies itself. During this process, the genetic code can randomly change in each copy. The changes are called mutations. If the coronavirus that causes COVID-19 changes in ways that make previous infections or vaccination less effective at preventing infection, people can get sick again.
The virus that causes COVID-19 can infect some pets. Cats, dogs, hamsters and ferrets have caught this coronavirus and had symptoms. It's rare for a person to get COVID-19 from a pet.
Risk factors
The main risk factors for COVID-19 are:
- If someone you live with has COVID-19 .
- If you spend time in places with poor air flow and a higher number of people when the virus is spreading.
- If you spend more than 30 minutes in close contact with someone who has COVID-19 .
Many factors affect your risk of catching the virus that causes COVID-19 . How long you are in contact, if the space has good air flow and your activities all affect the risk. Also, if you or others wear masks, if someone has COVID-19 symptoms and how close you are affects your risk. Close contact includes sitting and talking next to one another, for example, or sharing a car or bedroom.
It seems to be rare for people to catch the virus that causes COVID-19 from an infected surface. While the virus is shed in waste, called stool, COVID-19 infection from places such as a public bathroom is not common.
Serious COVID-19 illness risk factors
Some people are at a higher risk of serious COVID-19 illness than others. This includes people age 65 and older as well as babies younger than 6 months. Those age groups have the highest risk of needing hospital care for COVID-19 .
Not every risk factor for serious COVID-19 illness is known. People of all ages who have no other medical issues have needed hospital care for COVID-19 .
Known risk factors for serious illness include people who have not gotten a COVID-19 vaccine. Serious illness also is a higher risk for people who have:
- Sickle cell disease or thalassemia.
- Serious heart diseases and possibly high blood pressure.
- Chronic kidney, liver or lung diseases.
People with dementia or Alzheimer's also are at higher risk, as are people with brain and nervous system conditions such as stroke. Smoking increases the risk of serious COVID-19 illness. And people with a body mass index in the overweight category or obese category may have a higher risk as well.
Other medical conditions that may raise the risk of serious illness from COVID-19 include:
- Cancer or a history of cancer.
- Type 1 or type 2 diabetes.
- Weakened immune system from solid organ transplants or bone marrow transplants, some medicines, or HIV .
This list is not complete. Factors linked to a health issue may raise the risk of serious COVID-19 illness too. Examples are a medical condition where people live in a group home, or lack of access to medical care. Also, people with more than one health issue, or people of older age who also have health issues have a higher chance of severe illness.
Related information
- COVID-19: Who's at higher risk of serious symptoms? - Related information COVID-19: Who's at higher risk of serious symptoms?
Complications
Complications of COVID-19 include long-term loss of taste and smell, skin rashes, and sores. The illness can cause trouble breathing or pneumonia. Medical issues a person already manages may get worse.
Complications of severe COVID-19 illness can include:
- Acute respiratory distress syndrome, when the body's organs do not get enough oxygen.
- Shock caused by the infection or heart problems.
- Overreaction of the immune system, called the inflammatory response.
- Blood clots.
- Kidney injury.
Post-COVID-19 syndrome
After a COVID-19 infection, some people report that symptoms continue for months, or they develop new symptoms. This syndrome has often been called long COVID, or post- COVID-19 . You might hear it called long haul COVID-19 , post-COVID conditions or PASC. That's short for post-acute sequelae of SARS -CoV-2.
Other infections, such as the flu and polio, can lead to long-term illness. But the virus that causes COVID-19 has only been studied since it began to spread in 2019. So, research into the specific effects of long-term COVID-19 symptoms continues.
Researchers do think that post- COVID-19 syndrome can happen after an illness of any severity.
Getting a COVID-19 vaccine may help prevent post- COVID-19 syndrome.
The Centers for Disease Control and Prevention (CDC) recommends a COVID-19 vaccine for everyone age 6 months and older. The COVID-19 vaccine can lower the risk of death or serious illness caused by COVID-19.
The COVID-19 vaccines available in the United States are:
2023-2024 Pfizer-BioNTech COVID-19 vaccine. This vaccine is available for people age 6 months and older.
Among people with a typical immune system:
- Children age 6 months up to age 4 years are up to date after three doses of a Pfizer-BioNTech COVID-19 vaccine.
- People age 5 and older are up to date after one Pfizer-BioNTech COVID-19 vaccine.
- For people who have not had a 2023-2024 COVID-19 vaccination, the CDC recommends getting an additional shot of that updated vaccine.
2023-2024 Moderna COVID-19 vaccine. This vaccine is available for people age 6 months and older.
- Children ages 6 months up to age 4 are up to date if they've had two doses of a Moderna COVID-19 vaccine.
- People age 5 and older are up to date with one Moderna COVID-19 vaccine.
2023-2024 Novavax COVID-19 vaccine. This vaccine is available for people age 12 years and older.
- People age 12 years and older are up to date if they've had two doses of a Novavax COVID-19 vaccine.
In general, people age 5 and older with typical immune systems can get any vaccine approved or authorized for their age. They usually don't need to get the same vaccine each time.
Some people should get all their vaccine doses from the same vaccine maker, including:
- Children ages 6 months to 4 years.
- People age 5 years and older with weakened immune systems.
- People age 12 and older who have had one shot of the Novavax vaccine should get the second Novavax shot in the two-dose series.
Talk to your healthcare professional if you have any questions about the vaccines for you or your child. Your healthcare team can help you if:
- The vaccine you or your child got earlier isn't available.
- You don't know which vaccine you or your child received.
- You or your child started a vaccine series but couldn't finish it due to side effects.
People with weakened immune systems
Your healthcare team may suggest added doses of COVID-19 vaccine if you have a moderately or seriously weakened immune system. The FDA has also authorized the monoclonal antibody pemivibart (Pemgarda) to prevent COVID-19 in some people with weakened immune systems.
Control the spread of infection
In addition to vaccination, there are other ways to stop the spread of the virus that causes COVID-19 .
If you are at a higher risk of serious illness, talk to your healthcare professional about how best to protect yourself. Know what to do if you get sick so you can quickly start treatment.
If you feel ill or have COVID-19 , stay home and away from others, including pets, if possible. Avoid sharing household items such as dishes or towels if you're sick.
In general, make it a habit to:
- Test for COVID-19 . If you have symptoms of COVID-19 test for the infection. Or test five days after you came in contact with the virus.
- Help from afar. Avoid close contact with anyone who is sick or has symptoms, if possible.
- Wash your hands. Wash your hands well and often with soap and water for at least 20 seconds. Or use an alcohol-based hand sanitizer with at least 60% alcohol.
- Cover your coughs and sneezes. Cough or sneeze into a tissue or your elbow. Then wash your hands.
- Clean and disinfect high-touch surfaces. For example, clean doorknobs, light switches, electronics and counters regularly.
Try to spread out in crowded public areas, especially in places with poor airflow. This is important if you have a higher risk of serious illness.
The CDC recommends that people wear a mask in indoor public spaces if you're in an area with a high number of people with COVID-19 in the hospital. They suggest wearing the most protective mask possible that you'll wear regularly, that fits well and is comfortable.
- COVID-19 vaccines: Get the facts - Related information COVID-19 vaccines: Get the facts
- Comparing the differences between COVID-19 vaccines - Related information Comparing the differences between COVID-19 vaccines
- Different types of COVID-19 vaccines: How they work - Related information Different types of COVID-19 vaccines: How they work
- Debunking COVID-19 myths - Related information Debunking COVID-19 myths
Travel and COVID-19
Travel brings people together from areas where illnesses may be at higher levels. Masks can help slow the spread of respiratory diseases in general, including COVID-19 . Masks help the most in places with low air flow and where you are in close contact with other people. Also, masks can help if the places you travel to or through have a high level of illness.
Masking is especially important if you or a companion have a high risk of serious illness from COVID-19 .
- Goldman L, et al., eds. COVID-19: Epidemiology, clinical manifestations, diagnosis, community prevention, and prognosis. In: Goldman-Cecil Medicine. 27th ed. Elsevier; 2024. https://www.clinicalkey.com. Accessed Dec. 17, 2023.
- Coronavirus disease 2019 (COVID-19) treatment guidelines. National Institutes of Health. https://www.covid19treatmentguidelines.nih.gov/. Accessed Dec. 18, 2023.
- AskMayoExpert. COVID-19: Testing, symptoms. Mayo Clinic; Nov. 2, 2023.
- Symptoms of COVID-19. Centers for Disease Control and Preventions. https://www.cdc.gov/coronavirus/2019-ncov/symptoms-testing/symptoms.html. Accessed Dec. 20, 2023.
- AskMayoExpert. COVID-19: Outpatient management. Mayo Clinic; Oct. 10, 2023.
- Morris SB, et al. Case series of multisystem inflammatory syndrome in adults associated with SARS-CoV-2 infection — United Kingdom and United States, March-August 2020. MMWR. Morbidity and Mortality Weekly Report 2020;69:1450. DOI: http://dx.doi.org/10.15585/mmwr.mm6940e1external icon.
- COVID-19 testing: What you need to know. Centers for Disease Control and Prevention. https://www.cdc.gov/coronavirus/2019-ncov/symptoms-testing/testing.html. Accessed Dec. 20, 2023.
- SARS-CoV-2 in animals. American Veterinary Medical Association. https://www.avma.org/resources-tools/one-health/covid-19/sars-cov-2-animals-including-pets. Accessed Jan. 17, 2024.
- Understanding exposure risk. Centers for Disease Control and Prevention. https://www.cdc.gov/coronavirus/2019-ncov/your-health/risks-exposure.html. Accessed Jan. 10, 2024.
- People with certain medical conditions. Centers for Disease Control and Prevention. https://www.cdc.gov/coronavirus/2019-ncov/need-extra-precautions/people-with-medical-conditions.html. Accessed Jan. 10, 2024.
- Factors that affect your risk of getting very sick from COVID-19. Centers for Disease Control and Prevention. https://www.cdc.gov/coronavirus/2019-ncov/your-health/risks-getting-very-sick.html. Accessed Jan. 10, 2024.
- Regan JJ, et al. Use of Updated COVID-19 Vaccines 2023-2024 Formula for Persons Aged ≥6 Months: Recommendations of the Advisory Committee on Immunization Practices—United States, September 2023. MMWR. Morbidity and Mortality Weekly Report 2023; 72:1140–1146. DOI: http://dx.doi.org/10.15585/mmwr.mm7242e1.
- Long COVID or post-COVID conditions. Centers for Disease Control and Prevention. https://www.cdc.gov/coronavirus/2019-ncov/long-term-effects/index.html. Accessed Jan. 10, 2024.
- Stay up to date with your vaccines. Centers for Disease Control and Prevention. https://www.cdc.gov/coronavirus/2019-ncov/vaccines/stay-up-to-date.html. Accessed Jan. 10, 2024.
- Interim clinical considerations for use of COVID-19 vaccines currently approved or authorized in the United States. Centers for Disease Control and Prevention. https://www.cdc.gov/vaccines/covid-19/clinical-considerations/covid-19-vaccines-us.html#CoV-19-vaccination. Accessed Jan. 10, 2024.
- Use and care of masks. Centers for Disease Control and Prevention. https://www.cdc.gov/coronavirus/2019-ncov/prevent-getting-sick/about-face-coverings.html. Accessed Jan. 10, 2024.
- How to protect yourself and others. Centers for Disease Control and Prevention. https://www.cdc.gov/coronavirus/2019-ncov/prevent-getting-sick/prevention.html. Accessed Jan. 10, 2024.
- People who are immunocompromised. Centers for Disease Control and Prevention. https://www.cdc.gov/coronavirus/2019-ncov/need-extra-precautions/people-who-are-immunocompromised.html. Accessed Jan. 10, 2024.
- Masking during travel. Centers for Disease Control and Prevention. https://wwwnc.cdc.gov/travel/page/masks. Accessed Jan. 10, 2024.
- AskMayoExpert. COVID-19: Testing. Mayo Clinic. 2023.
- COVID-19 test basics. U.S. Food and Drug Administration. https://www.fda.gov/consumers/consumer-updates/covid-19-test-basics. Accessed Jan. 11, 2024.
- At-home COVID-19 antigen tests — Take steps to reduce your risk of false negative results: FDA safety communication. U.S. Food and Drug Administration. https://www.fda.gov/medical-devices/safety-communications/home-covid-19-antigen-tests-take-steps-reduce-your-risk-false-negative-results-fda-safety. Accessed Jan. 11, 2024.
- Interim clinical considerations for COVID-19 treatment in outpatients. Centers for Disease Control and Prevention. https://www.cdc.gov/coronavirus/2019-ncov/hcp/clinical-care/outpatient-treatment-overview.html. Accessed Jan. 11, 2024.
- Know your treatment options for COVID-19. U.S. Food and Drug Administration. https://www.fda.gov/consumers/consumer-updates/know-your-treatment-options-covid-19. Accessed Jan. 11, 2024.
- AskMayoExpert. COVID:19 Drug regimens and other treatment options. Mayo Clinic. 2023.
- Preventing spread of respiratory viruses when you're sick. Centers for Disease Control and Prevention. https://www.cdc.gov/respiratory-viruses/prevention/precautions-when-sick.html. Accessed March 5, 2024.
- AskMayoExpert. COVID-19: Quarantine and isolation. Mayo Clinic. 2023.
- COVID-19 resource and information guide. National Alliance on Mental Illness. https://www.nami.org/Support-Education/NAMI-HelpLine/COVID-19-Information-and-Resources/COVID-19-Resource-and-Information-Guide. Accessed Jan. 11, 2024.
- COVID-19 overview and infection prevention and control priorities in non-U.S. healthcare settings. Centers for Disease Control and Prevention. https://www.cdc.gov/coronavirus/2019-ncov/hcp/non-us-settings/overview/index.html. Accessed Jan. 16, 2024.
- Kim AY, et al. COVID-19: Management in hospitalized adults. https://www.uptodate.com/contents/search. Accessed Jan. 17, 2024.
- O'Horo JC, et al. Outcomes of COVID-19 with the Mayo Clinic Model of Care and Research. Mayo Clinic Proceedings. 2021; doi:10.1016/j.mayocp.2020.12.006.
- At-home OTC COVID-19 diagnostic tests. U.S. Food and Drug Administration. https://www.fda.gov/medical-devices/coronavirus-covid-19-and-medical-devices/home-otc-covid-19-diagnostic-tests. Accessed Jan. 22, 2024.
- Emergency use authorizations for drugs and non-vaccine biological products. U.S. Food and Drug Association. https://www.fda.gov/drugs/emergency-preparedness-drugs/emergency-use-authorizations-drugs-and-non-vaccine-biological-products. Accessed March 25, 2024.
- Coronavirus infection by race
- COVID-19 and pets
- COVID-19 and vitamin D
- COVID-19 and your mental health
- COVID-19 drugs: Are there any that work?
- COVID-19 in babies and children
- COVID-19 travel advice
- COVID-19 vaccines
- COVID-19 vaccines for kids: What you need to know
- COVID-19 variant
- COVID-19 vs. flu: Similarities and differences
- COVID-19, cold, allergies and the flu
- COVID-19: How can I protect myself?
- COVID-19: Who's at higher risk of serious symptoms?
- Debunking coronavirus myths
- Different COVID-19 vaccines
- Fight coronavirus (COVID-19) transmission at home
- Herd immunity and coronavirus
- How do COVID-19 antibody tests differ from diagnostic tests?
- How well do face masks protect against COVID-19?
- Is hydroxychloroquine a treatment for COVID-19?
- Long-term effects of COVID-19
- Mayo Clinic Minute: How dirty are common surfaces?
- Mayo Clinic Minute: You're washing your hands all wrong
- Pregnancy and COVID-19
- Safe outdoor activities during the COVID-19 pandemic
- Safety tips for attending school during COVID-19
- Sex and COVID-19
- Treating COVID-19 at home
- Unusual symptoms of coronavirus
Associated Procedures
- Convalescent plasma therapy
- COVID-19 antibody testing
- COVID-19 tests
- Extracorporeal membrane oxygenation (ECMO)
News from Mayo Clinic
- A Mayo Clinic virologist explains FLiRT and why you may need a new COVID-19 vaccination May 30, 2024, 02:30 p.m. CDT
- Mayo Clinic Q and A: Who should get the latest COVID-19 vaccine? Nov. 21, 2023, 01:30 p.m. CDT
- Can you get COVID-19 and the flu at the same time? A Mayo Clinic expert weighs in Oct. 16, 2023, 04:30 p.m. CDT
- At-home COVID-19 tests: A Mayo Clinic expert answers questions on expiration dates and the new variants Sept. 18, 2023, 04:00 p.m. CDT
- Mayo Clinic expert answers questions about the new COVID-19 vaccine Sept. 13, 2023, 04:15 p.m. CDT
- Study identifies risk factors for long-haul COVID disease in adults Sept. 13, 2023, 02:00 p.m. CDT
- Mayo researchers find vaccine may reduce severity of long-haul COVID symptoms Aug. 23, 2023, 04:34 p.m. CDT
- Corticosteroids lower the likelihood of in-hospital mortality from COVID-19 Aug. 04, 2023, 03:00 p.m. CDT
- COVID-19 vaccine administration simplified April 21, 2023, 07:00 p.m. CDT
- Science Saturday: COVID-19 -- the pandemic that's forever changed laboratory testing April 15, 2023, 11:00 a.m. CDT
- Mayo Clinic expert talks about the new omicron variant April 13, 2023, 02:13 p.m. CDT
- Mayo Clinic to ease universal face mask requirement April 04, 2023, 03:05 p.m. CDT
- 'Deaths of Despair' contribute to 17% rise in Minnesota's death rate during COVID-19 pandemic March 13, 2023, 12:00 p.m. CDT
- Rising cases of COVID-19 variant, XBB.1.5 Jan. 09, 2023, 05:15 p.m. CDT
- Bivalent COVID-19 booster approved for children 6 months and older Dec. 09, 2022, 09:33 p.m. CDT
- Mayo Clinic Minute: How to self-care at home when you have COVID-19 Dec. 06, 2022, 05:00 p.m. CDT
- Halloween safety tips from a Mayo Clinic infectious diseases expert Oct. 27, 2022, 02:00 p.m. CDT
- COVID-19, RSV and flu--season of respiratory infections Oct. 26, 2022, 04:30 p.m. CDT
- COVID-19 bivalent booster vaccines for kids 5-11 approved, Mayo Clinic awaits supply Oct. 13, 2022, 04:54 p.m. CDT
- Questions answered about the COVID-19 bivalent booster vaccines Oct. 12, 2022, 03:30 p.m. CDT
- Will the COVID-19 booster be like an annual flu shot? Sept. 12, 2022, 04:30 p.m. CDT
- Mayo Clinic Q and A: Who needs back-to-school COVID-19 vaccinations and boosters? Sept. 04, 2022, 11:00 a.m. CDT
- Q&A podcast: Updated COVID-19 boosters target omicron variants Sept. 02, 2022, 12:30 p.m. CDT
- Mayo Clinic Minute: Back-to-school COVID-19 vaccinations for kids Aug. 15, 2022, 03:15 p.m. CDT
- Mayo Clinic research shows bebtelovimab to be a reliable option for treating COVID-19 in era of BA.2, other subvariants Aug. 15, 2022, 02:09 p.m. CDT
- Mayo Clinic Q and A: New variants of COVID-19 Aug. 04, 2022, 12:30 p.m. CDT
- COVID-19 variant BA.5 is dominant strain; BA.2.75 is being monitored July 28, 2022, 02:30 p.m. CDT
- Mayo Clinic researchers pinpoint genetic variations that might sway course of COVID-19 July 25, 2022, 02:00 p.m. CDT
Products & Services
- A Book: Endemic - A Post-Pandemic Playbook
- A Book: Future Care
- Begin Exploring Women's Health Solutions at Mayo Clinic Store
- Symptoms & causes
- Diagnosis & treatment
- Doctors & departments
- COVID-19 vaccines: Get the facts
- How well do face masks protect against coronavirus?
- Post-COVID Recovery
News on coronavirus disease 2019 (COVID-19)
Learn the latest medical news about COVID-19 on Mayo Clinic News Network.
Double your impact on fighting cancer
Make a gift before July 31 and it can go twice as far to fight cancer.
Overview of Viral Infections
- Diagnosis |
- Treatment |
- Prevention |
A virus is composed of nucleic acid, either DNA or RNA , surrounded by a protein coat. It requires a living cell in which to multiply. A viral infection can lead to a spectrum of symptoms from asymptomatic (no overt symptoms) to severe disease.
People may get viruses by swallowing or inhaling them, by being bitten by insects, through sexual contact, or congenitally (passed by a pregnant person to the fetus).
Most commonly, viral infections involve the nose, throat, and upper airways, or systems such as the nervous, gastrointestinal, and reproductive systems.
Doctors may base the diagnosis on symptoms, blood tests and cultures, or examination of infected tissues.
Antiviral drugs may interfere with the reproduction of viruses or strengthen the immune response to the viral infection.
A virus is a small infectious organism—much smaller than a fungus or bacterium—that must invade a living cell to reproduce (replicate). The virus attaches to a cell (called the host cell), enters the cell, and releases its DNA or RNA inside the cell. The virus’s DNA or RNA is the genetic material containing the information needed to replicate the virus. The virus’s genetic material takes control of the host cell and forces it to replicate the virus. The infected cell usually dies because the virus keeps it from performing its normal functions. When the infected host cell dies, it releases new viruses, which go on to infect other cells.
Viruses are classified as DNA viruses or RNA viruses, depending on whether they use DNA or RNA to replicate. DNA viruses include herpesviruses . RNA viruses include SARS-CoV2 , which causes COVID-19. RNA viruses also include retroviruses, such as HIV ( human immunodeficiency virus ). RNA viruses, particularly retroviruses, are prone to mutate, meaning the set of genetic instructions that contain all the information that the virus needs to function can change as the virus spreads.
Some viruses do not kill the cells they infect but instead alter the cell's functions. Sometimes the infected cell loses control over normal cell division and becomes cancerous.
Some viruses, such as hepatitis B virus and hepatitis C virus , can cause chronic infections. Chronic hepatitis can last for years, even decades. In many people, chronic hepatitis is quite mild and causes little liver damage. However, in some people, it eventually results in cirrhosis (severe scarring of the liver), liver failure , and sometimes liver cancer .
Did You Know...
|
Viruses usually infect one particular type of cell. For example, common cold viruses infect only cells of the upper respiratory tract. Additionally, most viruses infect only a few species of plants or animals. Some infect only people.
Many viruses commonly cause infections in infants and children and older adults.
Types of viral infections
Upper respiratory infections (infections of the nose, throat, upper airways, and lungs) are likely the most common viral infections.
Upper respiratory infections include sore throat , sinusitis , and the common cold . Other viral respiratory infections include influenza , pneumonia , and coronaviruses , including SARS-CoV-2 (the virus that causes COVID-19).
In small children, viruses also commonly cause croup (which is inflammation of the upper and lower airways, called laryngotracheobronchitis) or lower airways ( bronchiolitis ).
Respiratory infections are more likely to cause severe symptoms in infants, older people, and people with a lung or heart disorder. Respiratory viruses are typically spread from person to person by contact with infected respiratory droplets.
Other viruses infect other specific parts of the body:
Gastrointestinal tract: Infections of the gastrointestinal tract, such as gastroenteritis , are commonly caused by viruses, such as noroviruses and rotaviruses .
Liver: These infections result in hepatitis .
Nervous system : Some viruses, such as the rabies virus and the West Nile virus , infect the brain, causing encephalitis. Others infect the layers of tissue that cover the brain and spinal cord (meninges), causing meningitis .
Skin: Viral infections that affect only the skin sometimes result in warts or other blemishes. Many viruses that affect other parts of the body, such as chickenpox , also cause a rash.
Placenta and fetus: Some viruses, such as the Zika virus , the rubella virus, and cytomegalovirus , can infect the placenta and fetus in pregnant women.
Some viruses typically affect many body systems. Such viruses include enteroviruses (such as coxsackieviruses and echoviruses) and cytomegaloviruses.
Spread of viruses
Viruses are spread (transmitted) in various ways. They may be
Spread by the bites of insects, such as mosquitoes, certain biting flies, or ticks
Spread sexually (in sexually transmitted infections )
Spread during transfusion of contaminated blood
Spread congenitally during pregnancy
New human viruses sometimes develop from viruses that usually affect animals (for example, SARS-CoV and SARS-CoV-2 ). This happens when the infected animal host comes into close contact with susceptible humans.
Many viruses that were once present in only a few parts of the world are now spreading. These viruses include chikungunya virus, Crimean-Congo hemorrhagic fever virus, Japanese encephalitis virus, Rift Valley Fever virus, West Nile virus , Ross River virus, Zika virus , and louping ill virus. These viruses are spreading partly because climate change has resulted in more areas where the mosquitoes or ticks that spread the viruses can live. Also, travelers may be infected, then return home and be bitten by a mosquito, which spreads the virus to other people.
Defenses against viruses
The body has a number of defenses against viruses:
Physical barriers, such as the skin, which discourage easy entry
The body's immune defenses, which attack the virus
When a virus enters the body, it triggers the body's immune defenses. These defenses begin with white blood cells , such as lymphocytes and monocytes, which learn to attack and destroy the virus or the cells the virus has infected. If the body survives the virus attack, some of the white blood cells remember the invader and are able to respond more quickly and effectively to a subsequent infection by the same virus. This response is called immunity. Immunity can also be produced by getting a vaccine .
Viruses and cancer
Some viruses alter the DNA of their host cells in a way that helps cancer develop. Some viruses, such as herpesviruses and HIV , leave their genetic material in the host cell, where the material remains dormant for an extended time (called latent infection). When the cell is disturbed, the virus may begin replicating again and cause disease.
Only a few viruses are known to cause cancer, but there may be others.
Diagnosis of Viral Infections
A doctor's evaluation
For infections that occur in epidemics, the presence of other similar cases
For some infections, blood tests and cultures
Common viral infections (such as measles , rubella , or chickenpox ) may be diagnosed based on symptoms.
For infections that occur in epidemics (such as influenza ), the presence of other similar cases may help doctors identify a particular infection. Laboratory diagnosis is important for distinguishing between different viruses that cause similar symptoms, such as COVID-19 (SARS-CoV2) and influenza .
For other infections, blood tests and cultures (growing microorganisms in the laboratory from samples of blood, body fluid, or other material taken from an infected area) may be done. Polymerase chain reaction (PCR) techniques may be used to make many copies of the viral genetic material. PCR techniques make it easier for doctors to rapidly and accurately identify the virus. Blood may also be tested for antigens, which are proteins on or in viruses that trigger the body's defense. Blood may also be tested for antibodies to viruses. (Antibodies are proteins produced by the immune system to help defend the body against a particular attack.) Tests are usually done quickly, especially when the infection is a serious threat to public health or when symptoms are severe.
A sample of blood or other tissues is sometimes examined with an electron microscope, which provides high magnification with clear resolution.
Treatment of Viral Infections
Treatment of symptoms.
Sometimes antiviral drugs
There are no specific treatments for many viruses. However, many things can help relieve certain symptoms, such as the following:
Dehydration: Plenty of fluids, sometimes given by vein (intravenously)
nonsteroidal anti-inflammatory drugs (NSAIDs)
Some rashes: Soothing or moisturizing creams and sometimes an antihistamine taken by mouth for itching
Not everyone who has these symptoms needs treatment. If symptoms are mild, it may be better to wait for them to go away on their own. Some treatments may not be appropriate for infants and young children.
Antiviral drugs
Medications that combat viral infections are called antiviral drugs. Many viral infections do not have effective antiviral drugs available to treat them. However, there are several drugs for influenza , many drugs for infection by one or more herpesviruses (see table Some Antiviral Drugs for Herpesvirus Infections ), and many antiviral drugs for treatment of HIV , hepatitis C , hepatitis B , and COVID-19 , which is caused by SAR-CoV-2.
Many antiviral drugs work by interfering with replication of viruses. Most drugs used to treat HIV infection work this way. Because viruses are tiny and replicate inside cells using the cells' own metabolic functions, there are only a limited number of metabolic functions that antiviral drugs can target. In contrast, bacteria are relatively large organisms, commonly reproduce by themselves outside of cells, and have many metabolic functions that antibacterial drugs (antibiotics) can target. Therefore, antiviral drugs are much more difficult to develop than antibiotics. Also, unlike antibiotics, which are usually effective against many different species of bacteria, most antiviral drugs are usually effective against only one (or a very few) viruses.
Antiviral drugs can be toxic to human cells. Also, viruses can develop resistance to antiviral drugs.
Most antiviral drugs can be given by mouth. Some can also be given by injection into a vein (intravenously) or muscle (intramuscularly). Some are applied as ointments, creams, or eye drops or are inhaled as a powder.
Antibiotics are not effective against viral infections, but if a person has a bacterial infection in addition to a viral infection, an antibiotic is often necessary.
Interferons are replicas of naturally occurring substances that slow or stop viral replication. These drugs are used to treat certain viral infections such as
Chronic hepatitis B
Chronic hepatitis C
Genital warts
Interferons may have side effects, such as fever, chills, weakness, and muscle aches. These effects typically start 7 to 12 hours after the first injection and last up to 12 hours.
Antibodies from the blood of a person who has recovered from the viral infection (convalescent serum) and antibodies that are produced in a laboratory from living cells that have been altered to produce the desired antibodies ( monoclonal antibodies ) are used to treat some viral infections including (for example, respiratory syncytial virus [RSV] infection , rabies ).
Prevention of Viral Infections
Prevention of viral infections may include
General measures
Immune globulins.
Vaccines and immune globulins help the body better defend itself against diseases caused by certain viruses (or bacteria). The process of strengthening the body's defenses is called immunization .
People can help prevent many viral infections by routine measures to protect themselves and others (personal protective measures). These measures vary depending on the how the virus is spread. Measures include the following:
Frequently and thoroughly washing the hands with soap
Consuming only food and liquids that have been appropriately prepared or treated
Avoiding contact with infected people and contaminated surfaces
Sneezing and coughing into tissues (which should be thrown away) or into the upper arm, completely covering the mouth and nose
Using safe-sex practices
Preventing bites by ticks , mosquitoes, and other arthropods
Mask wearing
Physical distancing when appropriate (for example, for COVID-19 prevention )
Vaccines work by stimulating the body’s natural defense mechanisms (called active immunization ). Vaccines are given before exposure to a virus to prevent infection.
Viral vaccines in general use include the following:
Hepatitis A
Hepatitis B
Human papillomavirus (HPV)
Japanese encephalitis (inflammation of the brain)
Measles, mumps, and rubella
Shingles ( herpes zoster )
Yellow fever
Adenovirus, smallpox , and mpox vaccines are available but used only in people who are at high risk of getting the infection, such as certain military personnel.
Viral diseases can be eradicated by effective vaccines. Smallpox was eradicated in 1978. Extensive vaccination has almost eradicated polio worldwide, but cases still occur in areas with incomplete immunization, such as sub-Saharan Africa and southern Asia. Measles has been almost eradicated from some parts of the world, such as the Americas. However, because measles is highly contagious and vaccination coverage is incomplete even in regions where it is considered eradicated, it is not likely to be completely eliminated soon.
Immune globulins are a sterilized solution of antibodies (also called immunoglobulins) collected from the blood of a group of people. Immune globulins are given directly to a person (called passive immunization) .
Immunoglobulins can be collected from the blood of the following:
People who are generally healthy (these immunoglobulins are called pooled human immunoglobulin)
People who have many antibodies that defend against a specific infectious organism, often because they have been infected with that organism (these immunoglobulins are called hyperimmune globulin)
Hyperimmune globulin is available for only a few infectious diseases, such as hepatitis B , rabies , tetanus , and chickenpox . It is usually given after people have been exposed to a microorganism but before they get sick. For example, people who have been bitten by an animal that might have rabies are immediately given rabies hyperimmune globulin.
Immune globulins are given by injection into a muscle or into a vein. The immunity provided by immune globulins lasts for only a few days or weeks, until the body eliminates the injected antibodies.
Sometimes, such as when people are exposed to rabies or hepatitis B, they are given both immune globulin and a vaccine to help prevent infection from developing or reduce the severity of infection.
Immune globulins may also help treat some infections. For example, they may be given to people whose immune system does not respond adequately to an infection (see Replacing missing parts of the immune system ).

Got any suggestions?
We want to hear from you! Send us a message and help improve Slidesgo
Top searches
Trending searches

stop bullying
11 templates

44 templates

welcome back
90 templates

27 templates

business pitch
695 templates
Infectious Disease
It seems that you like this template, infectious disease presentation, free google slides theme, powerpoint template, and canva presentation template.
Any disease caused by bacteria or viruses invading our body tissues are called infectious diseases. As always, every single bit of information that you can share about this topic to the medical community is welcome. With this template, you'll get a dark-colored design with some irregular shapes that have gradients. There are some layouts that you could use in your presentation, for example, to talk about risk factors, prevention or treatment.
Features of this template
- 100% editable and easy to modify
- 28 different slides to impress your audience
- Contains easy-to-edit graphics such as graphs, maps, tables, timelines and mockups
- Includes 500+ icons and Flaticon’s extension for customizing your slides
- Designed to be used in Google Slides, Canva, and Microsoft PowerPoint
- 16:9 widescreen format suitable for all types of screens
- Includes information about fonts, colors, and credits of the free resources used
How can I use the template?
Am I free to use the templates?
How to attribute?
Attribution required If you are a free user, you must attribute Slidesgo by keeping the slide where the credits appear. How to attribute?

Register for free and start downloading now
Related posts on our blog.

How to Add, Duplicate, Move, Delete or Hide Slides in Google Slides

How to Change Layouts in PowerPoint

How to Change the Slide Size in Google Slides
Related presentations.

Premium template
Unlock this template and gain unlimited access


An official website of the United States government
The .gov means it’s official. Federal government websites often end in .gov or .mil. Before sharing sensitive information, make sure you’re on a federal government site.
The site is secure. The https:// ensures that you are connecting to the official website and that any information you provide is encrypted and transmitted securely.
- Publications
- Account settings
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now .
- Advanced Search
- Journal List
- Taylor and Francis - PMC COVID-19 Collection
Epidemiology, pathogenesis, clinical presentations, diagnosis and treatment of COVID-19: a review of current evidence
Sayeeda rahman.
School of Medicine, American University of Integrative Sciences (AUIS), Bridgetown, Barbados
Maria Teresa Villagomez Montero
Kherie rowe, rita kirton, frank kunik, jr, introduction.
The COVID-19 pandemic has created a public health crisis, infected millions of people, and caused a significant number of deaths. SARS-CoV-2 transmits from person to person through several routes, mainly via respiratory droplets, which makes it difficult to contain its spread into the community. Here, we provide an overview of the epidemiology, pathogenesis, clinical presentation, diagnosis, and treatment of COVID-19.
Areas covered
Direct person-to-person respiratory transmission has rapidly amplified the spread of coronavirus. In the absence of any clinically proven treatment options, the current clinical management of COVID-19 includes symptom management, infection prevention and control measures, optimized supportive care, and intensive care support in severe or critical illness. Developing an effective vaccine is now a leading research priority. Some vaccines have already been approved by the regulatory authorities for the prevention of COVID-19.
Expert opinion
General prevention and protection measures regarding the containment and management of the second or third waves are necessary to minimize the risk of infection. Until now, four vaccines reported variable efficacies of between 62–95%, and two of them (Pfizer/BioNTech and Moderna) received FDA emergency use authorization. Equitable access and effective distribution of these vaccines in all countries will save millions of lives.
1. Introduction
Coronavirus disease 2019 (COVID-19) is a highly contagious and infectious disease caused by the novel coronavirus, severe acute respiratory syndrome Coronavirus-2 (SARS-CoV-2) [ 1 , 2 ]. It is well documented that the initial cases of COVID-19 related infection were first reported in Wuhan, Hubei Province of China in December 2019, and were linked to the Huanan Seafood Market [ 3 ]. Since then, the infection has spread to over 216 countries and territories. The World Health Organization (WHO) announced that COVID-19 reached pandemic status on 30 January 2020 [ 4 , 5 ] and subsequently, declared a global pandemic in March 2020 [ 6 ]. It has since been referred to be ‘the most crucial global health calamity of the century and the greatest challenge that humankind faced since the 2nd World War’ [ 7 ]. As of 26 December 2020, there were approximately 80,500,000 confirmed COVID-19 cases worldwide, including 1,700,000 related deaths [ 8 ], with a case fatality rate of 2.2%. The case fatality rate varies among countries, estimated from 0 to more than 20% [ 9 ]. A second wave of COVID-19 infection has already been recorded in many countries, which may be due to premature relaxation of government-enforced lockdown rules in many parts of the world [ 10 , 11 ]. Several countries have reported a new rise in daily cases higher than the first wave in March 2020 [ 12 , 13 ]. Although there is no shortage of information on this pandemic virus presented in everyday practice, this paper presents a comprehensive review of the latest information on SARS-CoV-2 highlighting the epidemiology, pathogenesis, and clinical aspects of SARS-CoV-2 infection.
We searched and reviewed literature published since November 2019, which focused on the epidemiology, pathogenesis, diagnosis, treatment, and prevention of COVID-19. Original studies, reviews, editorials, commentaries, perspectives, short or special communications, and position/policy papers on the COVID-19 pandemic were also searched. Information from websites of different professional associations and national or international organizations was extracted. Reference lists from the retrieved articles were also manually examined for relevant information. PubMed, Scopus, and Google Scholar were also searched using specific keywords, including ‘SARS-CoV-2ʹ, ‘COVID-19 infection’, ‘epidemiology’, ‘pathogenesis’, ‘diagnosis’, ‘treatment’, and ‘prevention’.
3. Origin, history, and epidemiology of COVID-19
Coronaviruses are a large family of viruses that are common in humans and many different species of animals (e.g. cats, bats). Most people are infected with these viruses at some point in their lives. Common human coronaviruses typically cause upper respiratory tract infections (URTIs) such as the common cold. However, some variants can cause mild influenza-like symptoms. Initially, cases related to SARS-CoV-2 were associated with high mortality rates, especially in people with chronic diseases, such as diabetes and cardiovascular diseases [ 14 , 15 ].
There are four main genres of coronaviruses: alpha (α), beta (β), gamma (γ), and delta (δ). The first human coronaviruses were identified in the mid-1960s. Common variants that affect people around the world include 229E, NL63, OC43, and HKU1. Among them, 229E and NL63 are α-coronaviruses, and OC43 and HKU1 are β-coronaviruses [ 16 ]. The usual signs and symptoms generated by these coronaviruses are similar to those of the common cold, accompanied by mild to moderate URTI. It is also of note that some coronaviruses that infect animals can undergo mutation and adaptation, thereby driving the co-evolution of coronaviruses that can become a new human coronavirus (HCoV) [ 17 ]. Therefore, these HCoV infections are zoonotic, and their symptoms are accompanied by more severe respiratory tract syndromes than those of the aforementioned ones. Three recent examples of these are: (i) SARS-CoV-2 (the novel coronavirus, causing coronavirus disease in 2019 or COVID-19), (ii) SARS-CoV (the β-coronavirus, causing severe acute respiratory syndrome, or SARS), and (iii) MERS-CoV (the β-coronavirus, causing Middle East respiratory syndrome, or MERS) [ 17 , 18 ].
COVID-19 was initially thought to be a zoonotic disease originating in bats, which may have undergone several cross-species events, first crossing the species barrier to pangolins and subsequently to humans. The outbreak appeared to have started from single or multiple zoonotic transmission events in the wet market in Wuhan [ 19 ]. As such, it was initially suspected that direct contact with intermediate host animals or the consumption of wild animals was the main route of SARS-CoV-2 transmission [ 5 ]. Its epidemiological link was first demonstrated by the appearance of several reported cases of severe respiratory distress, which had a typical characteristic radiological pattern (e.g. initial chest images demonstrated multifocal airspace opacities and consolidation in 70–80% of coronavirus-infected patients [ 20 ]). SARS-CoV-2 is highly transmissible and preliminary reports have suggested that the reproductive number (R 0 ) of people that an infected person could potentially infect is approximately 2.2 [ 21 ]. The R 0 is used to reflect contagious disease, and the higher the number, the more infectious the disease. If SARS-CoV-2 is compared to influenza and other diseases, the high R 0 , which varies from to 3–5, is representative of a more contagious infection [ 22 , 23 ] ( Table 1 ). The number of COVID-19 cases increased at a rapid rate, partly due to the highly infectious nature of the virus as well as the lack of awareness and availability of diagnostic kits in the initial stages of the pandemic [ 24 ].
Reproduction number (R 0 ) of some selected viruses [ 22 , 23 ]
| Viruses | R |
|---|---|
| Measles | 12–18 |
| Pertussis (Whooping cough) | 12–17 |
| Chickenpox | 8–9 |
| Rhinovirus (cold) | 5–7 |
| COVID-19 | 3–5 |
| Smallpox | 5–7 |
| HIV/AIDS | 2–5 |
| SARS | 2–5 |
| 1918 influenza | 2–3 |
| Seasonal influenza | 1–2 |
Mortality for COVID-19 appears to be higher than that for influenza, especially seasonal influenza. Early estimates relied heavily on genetic tests, which are the gold standard for diagnosing COVID-19, from either sputum or nose swabs from the back of the nose [ 25 ]. However, these tests only provide a clear picture of active infection; they are not an accurate reflection of possible past infective events. In addition to the genetic tests, serological studies are now also used, and can indicate whether the individual has been infected in the past, based on antibody response [ 26 ].
4. Structural and molecular features of SARS-CoV-2
SARS-CoV-2 belongs to the genus Betacoronavirus of the subfamily Orthocoronavirinae in the family Coronaviridae, and the order Nidovirales [ 27–30 ]. The viral particle is pleomorphic, as confirmed by cryo-electron tomography, and possesses non-segmented, single-stranded, positive-sense ribonucleic acid (ssRNA+) as its genome [ 31 , 32 ]. A coronavirus contains four structural base proteins: spike (S), envelope (E), membrane (M), and nucleocapsid (N) [ 32 , 33 ]. Among them, the S protein plays the most important role in viral attachment, fusion, and entry [ 34 ]. Its 30 kb genome RNA is large enough to produce a positive sense to be read directly by ribosomes in the cell [ 33 ]. The genome is coated with an N protein, which forms a helical nucleocapsid [ 35 ]. The N protein-coated genome is enclosed in a lipid envelope, and the viral lipid envelope is speckled by viral proteins [ 35 , 36 ]. As viruses cannot make their own lipids, they use the host’s lipids for replication and morphogenesis [ 37 ]. The N protein plays a crucial role in the morphogenesis phase of the viral life cycle during virion formation [ 35 ]. In addition to the lipid envelope, coronaviruses have a membrane glycoprotein called the matrix protein on its outer layer [ 38 ]. This transmembrane protein has a significant C-terminal domain that makes contact with the N protein [ 39 ]. Another minor envelope protein, E, is also an important component at the end of the viral life cycle [ 38 ].
Coronaviruses get their name from the characteristic feature of their S protein, which resembles a halo effect seen in solar eclipse or a crown-like appearance under an electron microscope [ 34 ]. The S protein has a roughly cylindrical shape and is heavily glycosylated [ 40 ], and encodes and possesses both receptor-binding and-fusion functions. Coronavirus uses its S protein, a main target for neutralizing antibodies, to bind with specific receptors and mediate membrane fusion and virus entry. It is a trimeric protein [ 34 ], composed of three intertwined chains that have identical amino acid sequences, each of which is called a protomer. However, the protomers do not have identical three-dimensional conformations. The monomer of the trimeric S protein is approximately 180 kDa and contains two distinct functional subunits, S1 and S2, both necessary for mediating attachment and membrane fusion, respectively. In its structure, N- and C-terminal portions of the S1 fold are two independent domains, the N-terminal domain (NTD) and C-terminal domain (CTD). Depending on the virus, either NTD or CTD can serve as the receptor-binding domain (RBD). The S protein induces successful infusion into the cell by first binding to the host receptor through the RBD of the S1 subunit, resulting in viral genomic fusion; the second stage by S2 facilitates the fusion of the cell and host membranes, which contains amino acid sequences necessary for continuing infiltration [ 41–43 ]. The RBD in the S protein is the most mutable part of the coronavirus genome and tends to be common for general viruses [ 44 ].
During viral replication, SARS-CoV-2 uses host protease enzymes to covalently attach sugars to asparagine side chains near the protein surface [ 45 ]. To achieve fusion, the S protein needs to be cleaved by proteases present in the host cell. The hosts own peptide bond breaking proteases cut the S protein at specific sites, and conformational changes enable fusion to occur [ 46 ]. Moreover, the availability of proteases on target cells largely determines whether coronaviruses enter cells through the plasma membrane or by endocytosis [ 47 ]. Proteolytic cleavage of the S glycoprotein also determines whether the virus can cross species, for example, from bats to humans [ 48 ]. The process is critical because it allows the fusion sequences to be exposed. The nature of the cell protease that cleaves the S glycoprotein varies according to the coronavirus [ 31 ]. Coronavirus proteins may be cleaved by one or several host proteases based on virus strains and cell types, including trypsin, cathepsins, transmembrane protease serine protease-2 (TMPRSS-2), TMPRSS-4, or human airway trypsin-like protease (HAT) [ 43 , 49 ]. However, the specific proteases that promote virus entry into SARS-CoV-2 remain elusive [ 43 , 49 , 50 ]. This cleavage is generally mediated by furin [ 50 ], an enzyme belonging to the subtilisin-like proprotein convertase family. It cleaves precursor proteins and facilitates their conversion to a biologically active state; thus, it plays a vital role in viral protein processing [ 51 ]. The S1/S2 cleavage site is the target site of furin during infection. The RBD of the S1 subunit contacts angiotensin-converting enzyme 2 (ACE2), which is facilitated by furin cleavage [ 52 , 53 ]. Furin proteases are found in significant amounts in the lungs. Therefore, viruses that attack the respiratory tract make use of this enzyme to convert and activate their own surface glycoproteins. Basically, it is like a lock-and-key mechanism, where viral glycoprotein and cellular receptor represent key and lock, respectively. Other influenza pathogens that have similar cleavage sites can also be acted upon by furin and other cellular proteases. The prevalent expression of cellular proteases across cell types increases the potential for the virus to successfully infiltrate the host [ 53 ]. It should be mentioned here that all other β-coronaviruses, including SARS-CoV, which is the closest to the SARS-CoV-2 strain, do not contain this cleavage site [ 54 ]. A study showed that the S protein of SARS-CoV-2 is 10 to 20 times more likely to bind to human ACE2 than the S protein of the early 2000s SARS-CoV strain [ 55 ]. The heightened affinity for a prevalent cellular receptor may be a factor that increases transmission [ 56 ].
5. Mechanism of SARS-Cov-2 transmission
5.1. mechanisms of transmission.
The transmissibility of an infection is determined by the basic R 0 , with a value above the threshold of 1 implies continuous and sustained human-to-human transmission [ 23 , 57 ]. The rapid spread of SARS-CoV-2 is due, in part, to the transmission mechanisms of the viral agent. An understanding of the transmission dynamics of infectious spread is critical, providing insights into the epidemiologic spread, implementation of outbreak control measures, and determination of the efficacy of such control measures [ 23 ].
The transmission characteristics of SARS-CoV-2 are very similar to those of SARS-CoV and pandemic influenza. Riou et al . [ 57 ] stated that this was an indicator of the potential for sustained human-to-human transmission and the risk of global spread. More recently, a mean R 0 range of 2.24 to 3.58 [ 58 , 59 ] was determined. With transmissibility on par with that of SARS-CoV, pandemic influenza, and HIV, but much lower than measles and chickenpox ( Table 1 ), SARS-CoV-2 presented a moderate to severe infectious threat [ 57 ].
The first evidence of potential person-to-person transmission was reported by Chan et al . [ 60 ]. They investigated the transmission of the virus in a group of family members who had recently visited Wuhan. They had no history of contact with animals, visits to markets, or eating game meat, but stayed in the same hotel throughout their travel. With no direct zoonotic involvement, this was the first indication that the virus could be spread by human contact. These initial findings were subsequently confirmed with increasing evidence demonstrating sustained human-to-human transmission [ 57 , 61 ].
SARS-CoV-2 uses the same receptor, ACE2, as SARS-CoV, and mainly spreads through the respiratory tract [ 62 ]. As a respiratory infectious disease, the virus is transmitted primarily by droplets, respiratory secretions, and direct contact [ 63 ]. However, viral particles have been isolated from fecal swabs and blood, implying several alternative routes for transmission [ 64–66 ]. It is worth noting that the ACE2 protein is also expressed by enterocytes in the small intestine [ 67 ]. Previous Chinese reports have shown no evidence of vertical transmission of the virus by blood products or the fecal-oral route [ 64 , 68–70 ]. However, some recent studies from the United Kingdom (UK) and other countries have confirmed a low rate of vertical transmission due to COVID-19 [ 71–75 ].
5.2. Incubation period
The incubation period on average is 1–14 days, however, generally is 3–7 days. SARS-CoV-2 may be present in the throat or the nose a few days before symptom onset. Interestingly, completely asymptomatic subjects may have viral loads similar to those of symptomatic patients [ 76 ]. This implies that asymptomatic individuals may be possible sources of infection. After the incubation period, patients present with similar symptoms, including fever, cough, and malaise. A small percentage of patients also manifest gastrointestinal symptoms, such as diarrhea and vomiting. The elderly and those with underlying disorders rapidly develop acute respiratory distress syndrome (ARDS), septic shock, metabolic acidosis, and coagulation dysfunction, which may ultimately lead to multiple organ failure and even death [ 5 , 77 , 78 ].
6. Clinical and pathological characteristics of COVID-19
SARS-CoV-2 targets the respiratory system, and transmission occurs via contact droplets and fomites from an infected person who may be symptomatic or asymptomatic [ 79 ]. During the incubation period, the virus triggers a slow response in the lungs. SARS-CoV-2 mainly invades alveolar epithelial cells, resulting in respiratory symptoms [ 80 ].
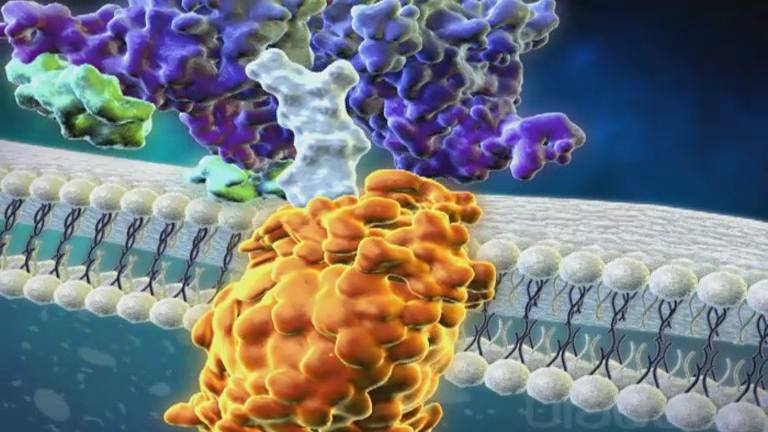
The S-glycoprotein on the surface of SARS-CoV-2 binds to ACE2 [ 80 ]. The receptor and the enzyme on the surface of type 2 alveolar cells induce a conformational change in S-glycoprotein initiating proteolytic digestion by host cell proteases (TMPRSS2 and furin), ultimately leading to internalization of the virion [ 81 ]. This implies that SARS-CoV-2 has a pathogenesis similar to that of SARS-CoV [ 82 ]. Coronaviruses generally enter via endocytosis or direct fusion of the viral envelope with the host membrane. Once internalized by the host cell, the viral particle is uncoated, and its genome enters the cell cytoplasm. Coronaviruses have an RNA genome from which they can directly produce their proteins and new genomes in the cytoplasm by attaching to the host ribosomes [ 83 ]. The host ribosomes translate viral RNA into RNA polymerase proteins. This RNA polymerase then reads the positive strand again to generate single-stranded, negative-sense RNA (ssRNA-) strands.
The ssRNA- strands are then used as a template by RNA polymerase to make additional ssRNA+ strands. The small RNA strands are read by host ribosomes in the endoplasmic reticulum to make the structural components of the virus. These structural components are then transferred from the endoplasmic reticulum to the Golgi apparatus. Within the Golgi apparatus, ssRNA+ genomes are packaged in the nucleocapsids to create new virion particles. These progeny viruses are then released from the host cell via exocytosis through secretory vesicles. The replication of the virus in alveolar cells mediates damage and induces an inflammatory response in the tissues. Cellular entry of the virus triggers an inflammatory response by recruiting T-helper cells that produce interferon (IFN)-gamma (IFN-γ), interleukin (IL)-2, and IL-12 [ 84 ]. The injured alveolar cells also release interferons, cytokines, and other intracellular components. The subsequent recruitment of other inflammatory cells leads to the development of a ‘ cytokine storm ’ which can precipitate the organ damage and multi-organ failure seen in severe disease [ 84 ]. COVID-19 infected patients have shown higher concentrations of peripheral blood immune mediators [ 85 ]. IL-6, interferon gamma-induced protein (IP)-10, and IFN-γ were markedly elevated in all three highly pathogenic HCoV infections [ 3 , 85 ]. Interferons act in a paracrine manner and can have numerous effects on the surrounding cells, preparing them against viral infection [ 86 ]. The alveolar macrophages detect cell injury and respond to cytokines released by injured alveolar cells. The alveolar macrophages respond by secreting cytokines and chemokines [ 87 ]. The inflammatory process occurring within the lung parenchyma stimulates nerve endings responsible for initiating the cough reflex, thus, people often present with an early dry cough [ 87 ]. Tumor necrosis factor (TNF)-α and IL-1β are proinflammatory cytokines that cause an increase in vascular permeability, increase in adhesion molecule expression, and induce recruitment of more immune cells, including neutrophils and monocytes. They bind to adhesion proteins on the surface of tissues and enter the site of injury [ 88 ]. IL-8 recruits neutrophils, and other chemokines attract monocytes [ 89 ]. The increase in vascular permeability causes leakage of fluid into the interstitial space and alveoli, resulting in interstitial and pulmonary edema. This can lead to dyspnea, impaired oxygenation, or hypoxemia. The clinicopathological characteristics of coronaviruses are shown in Figure 1 .

Viral replication of SARS-coV-2 in alveolar cells
Neutrophils engulf viruses and other debris around the area, which can be detrimental because this activity also results in the release of chemical by-products that damage the surrounding tissue [ 90 ]. Consequently, when there are damaged alveolar cells all over, less surfactant is produced. The alveoli can easily collapse, resulting in impaired oxygenation or hypoxemia [ 91 ] ( Figure 1 ). White blood cells (WBCs) and damaged endothelial cells release other inflammatory mediators, including arachidonic acid metabolites, including leukotrienes and prostaglandins. Leukotrienes cause bronchoconstriction, leading to impaired ventilation, and subsequent hypoxemia [ 92 ]. Prostaglandins, IL-1, IL-6 and TNF-α are responsible for causing fever, a primary feature of COVID-19 [ 93 , 94 ]. Decreased oxygen levels in the blood stimulate chemoreceptors in the cardiopulmonary center in the brain, which causes an increased inspiratory rate to increase oxygen levels in the blood and also initiate the heart to pump faster to deliver oxygen to the body [ 95 ]. For this, patients with hypoxemia usually develop tachypnea and tachycardia [ 96 ]. However, some patients may be asymptomatic because their immune system keeps it in check or only minor symptoms, such as cough accompanied by shortness of breath and some fever. The alveolar macrophages can also detect the virus using its special toll like receptor-4 (TLR-4) receptors, which engulf viral particles through phagocytosis [ 97 ].
A common finding in COVID-19 is lymphopenia, which is assumed to be due to the release of interferons [ 98 ]. IL-6 stimulates hepatocytes to produce acute phase reactants such as C-reactive protein (CRP), fibrinogen, and hepcidin [ 99 ]. CRP is a good inflammatory marker, and a high level in the blood is a marker of inflammation [ 100 ]. Therefore, the damaged alveolar tissue, accumulation of the fluid, ventilation/perfusion mismatch, and hypoxemia, which are not related to heart function, leads to the presentation of ARDS, which is considered to be the leading cause of mortality in COVID-19 [ 101 ].
6.1. Clinical manifestations
Patients with COVID-19 experience varying degrees of severity, and 80% of them have mild infection [ 102 ]. Approximately 15% of cases develop severe disease characterized by dyspnea, hypoxia, and lung changes on imaging; 5% are critically ill, with respiratory failure from ARDS, shock, and/or multi-organ dysfunction [ 3 , 103 , 104 ]. As ACE2 is expressed not only in the lungs but also in the heart, endothelium, renal tubular epithelium, intestinal epithelium, and the pancreas, SARS-CoV-2 may possess the potential to invade these tissues, to proliferate and destroy these organs, causing multiple organ dysfunction syndrome (MODS) [ 105 , 106 ]. Excessive activation of lymphocytes and increased pro-inflammatory mediators in patients with COVID-19 promotes immune-mediated damage. The process causes a mild disease to increase in severity and single organ involvement to progress to MODS. In severe cases, the disease can lead to ARDS, septic shock, metabolic acidosis, coagulation dysfunction, and MODS. Elderly individuals with reduced immunity and comorbidities are more susceptible to severe infections [ 107 ].
The median age of individuals affected by severe complications related to COVID-19 ranges from 49 to 56 years of age [ 108 ]. As symptoms progress, patients may develop pneumonia with ARDS, which requires intensive care. Children are typically asymptomatic or present with mild symptoms. Men and women have the same susceptibility to infection; however, male patients are more at risk for worse outcomes and death [ 109 ]. The symptoms include fever, fatigue, dry cough, anorexia, myalgia, dyspnea, and sputum production [ 110 ]. Mortality rate increases with age, with a significant increase above 80 years of age. The mortality rate also increased with comorbidities, including diabetes, heart disease, chronic kidney disease, chronic lung disease, and other socio-demographic factors ( Table 2 ). An increased risk of infection due to SARS-CoV-2 is also found to be associated with other comorbidities such as hypertension (27–30%), diabetes (19%), and coronary heart disease (6–8%) [ 104 , 111 ]. Studies have also demonstrated that patients with severe COVID-19 develop ARDS (67.3%), acute kidney injury (28.9%), abnormal hepatic function (28.9%), and cardiac injury (23.1%) [ 112 ]. An overview of the effect of COVID-19 on different pathophysiological conditions is presented in Table 2 [ 109 , 113–124 ].
Effect of COVID-19 on different pathophysiological conditions
| Sources | Pathophysiology | Pathogenesis of COVID-19 |
|---|---|---|
| Xu et al (2020) [ ] Gąsecka et al. (2020) [ ] | Respiratory diseases | of SARS-CoV-2 infection targets the nasal and bronchial epithelial cells and pneumocytes. of infection SARS-CoV-2 infects pulmonary capillary endothelial cells, accentuating inflammatory response and triggering an influx of monocytes and neutrophils [Ref]. , fulminant activation of coagulation and consumption of clotting factors occur indicated as diffuse intravascular coagulation. |
| Qian et al (2020) [ ] | Renal diseases | |
| Lippi et al (2020) [ ] | Hypertension | |
| Gamble et al (2020) [ ] Fang et al (2020) [ ] | Diabetes Mellitus | |
| Tham et al (2019) [ ] Memtsoudis et al (2020) [ ] Antonia et al (2020) [ ] | Obesity | |
| Vepa et al (2020) [ ] | Ethnicity | |
| Jin et al (2020) [ ] | Gender | |
| Rahman et al (2020) [ ] | Age | |
| Gérard et al (2020) [ ] | Blood group |
7. COVID-19 diagnostic techniques
The rapid and accurate detection of COVID-19 has become vital for effective response and prevention of further spread in large populations. Contact tracing has also been shown to be of extreme importance. It has allowed the systematic encapsulation of specific points of caseload increase, giving governments the opportunity to protect the health of the population without completely shutting down their economies. The American Center for Disease Control and Prevention (CDC) has been utilized since the initial identification of SARS-CoV-2 molecular assays for its detection, mostly using real-time polymerase chain reaction (PCR) methods [ 125 ]. The PCR for COVID-19 can only diagnose whether a person is currently infected with this particular coronavirus. It cannot provide information on other diseases or symptoms [ 126 ] and could miss patients who have cleared the virus and recovered from the disease [ 126 , 127 ]. Serology tests are also important as they can help assess the immune response [ 128 ], follow up on the progression of the disease, and the length of immune protection present after patients have recuperated from COVID-19 [ 129 ]. The serologic test is an enzyme-linked immunosorbent assay (ELISA)-based test that detects SARS-CoV-2 antibodies (IgG and IgM) in serum or plasma. The ELISA used by the CDC utilizes purified SARS-CoV-2 S protein (no live virus) as an antigen [ 130 ]. The problem with serologic tests is that the cross-reactivity to antibodies generated by other coronaviruses cannot be completely ruled out [ 130 ]. Comparative information on the use of different diagnostic techniques for COVID-19 is presented in Table 3 [ 131–134 ].
Viral test for COVID-19
| | | | | |||||
|---|---|---|---|---|---|---|---|---|
| Alcoba-Florez et al (2020) [ ] | Real-Time PCR(RT-PCR) | Viral RNA | Nasopharyngeal swab, sputum, stool | 3–4 hrs | Nucleic acid amplification test | - Gold standard diagnostic test. - Identifies directly the presence of virus. | Sensitive to sample collection error. Labor intensive. Specialized high-cost equipment. | >97%/>95% |
| Peto et al (2020) [ ] | LAMP | Viral RNA | Nasopharyngeal swab, sputum, stool | 2–3 hrs | Nucleic acid amplification test | - Cost-efficient. - Can be read by eye. | New techniques still under clinical investigation | >95% |
| Lisboa et al (2020) [ ] | ELISA | IgG or IgM | Blood | 1–3 hrs | Detection of IgM/IgG ot RBD IgG antibodies, via colorimetric assay | - Cost-effective. - Well documented in science. - Test 96 samples at a time. | Requires laboratory. Not well- established for SARS-CoV-2 | 79%/80% |
| Nicol et al (2020) [ ] | Lateral Flow Immunoassays | IgG or IgM | Blood | 15 to 20 min | Detection of IgM/IgG antibodies via color change of strip in lateral flow assay | - Extremely quick results. - Little training required. | - Evidence for accuracy still under investigation. - Expensive. - Not effective for large batch testing | 96%/80% |
8. Treatment and preventive measures
In the absence of any clinically proven treatment options, the treatment is symptomatic, and current clinical management includes infection prevention and control measures as well as supportive care [ 135 ]. Available therapeutic drugs include antiviral agents (e.g. remdesivir, hydroxychloroquine, chloroquine) and supporting agents (vitamin C, azithromycin, corticosteroids, IL-6 antagonists) [ 136 , 137 ]. Developing an effective COVID-19 vaccine is currently the world’s leading research priority [ 138 ]. Some vaccines have already been approved by the regulatory authorities for the prevention of COVID-19 [ 139–141 ].
8.1. Public health and preventive measures
Public health and preventive approaches are the current strategies to curb the transmission of COVID‐19 and focus on testing, case tracing, isolation, social distancing, and personal hygiene [ 142 ]. Important COVID-19 prevention and control measures in the community include hand hygiene, personal protective equipment (PPE), crowd avoidance, social distancing, isolation, school measures/closures, workplace measures/closures, quarantine, and travel restrictions [ 143 , 144 ].
A study conducted in Singapore recommended closing schools, maintaining effective social distancing in the workplace, and adopting quarantine measures to contain the pandemic once community transmission had been established [ 145 ]. Such measures were also found to reduce infection, mortality, and intensive care unit (ICU) admissions [ 58 , 146 , 147 ]. Social distancing reduces interactions between people and is effective in preventing community transmission [ 142 ]. The use of face masks is strongly indicated to reduce COVID-19 transmission in potentially asymptomatic or pre-symptomatic people [ 148 , 149 ]. The widespread use of face masks has been found to be effective in preventing SARS-CoV-2 transmission in highly affected areas in Italy and New York City [ 150 ]. Studies have demonstrated that a surgical mask could reduce virus exposure by an average of six times (range: 1.1 to 55 times) and should be worn by the potentially infected subject [ 151 ]. The WHO recommended the use of PPE by health care workers as they are more likely to be increasingly exposed to the virus and should wear medical/surgical masks, gowns, gloves, and face shields when treating infected patients or collecting samples [ 152 ].
Quarantine was found to be the most effective method for reducing the number of infected cases and decreasing mortality rates [ 22 , 153 , 154 ]. A review of 29 COVID-19 related studies found that quarantine can decrease the rate of infected cases (from 81% to 44%) and mortality (from 61% to 31%) [ 155 ]. Travel restrictions and lockdown in the early phase of the pandemic in Australia [ 156 ] and China [ 157 ] helped to decrease transmission effectively. Testing, isolation, and contact tracing were found to be effective in controlling the spread of the virus in countries such as South Korea, Singapore, Taiwan, and Hong Kong [ 158–161 ]. In contrast, Italy witnessed a wider outbreak as the country failed to employ such preventive measures during the early phase of the pandemic [ 161 ].
8.2. Management strategies based on symptoms
Management strategies of COVID-19 patients depend on the severity of the symptoms of the patients [ 162 , 163 ]:
- Mild cases:
- SpO 2 levels of 94%–97% in room air
- Symptomatic treatment
- O 2 therapy via nasal canula
- (2) Moderate cases:
- SpO 2 levels of 90%–94% in room air
- High-flow nasal oxygen (HFNO) therapy or noninvasive ventilation (NIV) in case of no improvement
- (3) Severe cases:
- SpO 2 levels ≤ 90% in room air or patients with ARDS
- O 2 therapy via HFNO/NIV with helmet
- Invasive ventilation via endotracheal intubation for patients with ARDS in cases of falling SpO 2 levels
- ARDS management
8.3. Pharmacological treatments
8.3.1. antiviral agents.
Extensive research is ongoing regarding antiviral therapies for the treatment of COVID-19. Although several antiviral therapies are being investigated by scientists, no treatments have been shown to be effective in treating COVID-19 [ 164 , 165 ]. Preliminary results are available from The Adaptive COVID-19 Treatment Trial (ACTT-1) from hospitalized COVID-19 patients. This double-blind randomized control trial (RCT) conducted in 60 trial sites and 13 subsites (United States of America [USA] (45 sites), Denmark (8), UK (5), Greece (4), Germany (3), Korea (2), Mexico (2), Spain (2), Japan (1), and Singapore (1)] showed that remdesivir was associated with a shorter median recovery time compared with placebo (11 vs. 15 days) with evidence of lower respiratory tract infection [ 165 ]. The trial also showed a significant mortality benefit (remdesivir group 4.0% vs. control group 12.7%). A study conducted in China, prematurely terminated due to adverse events of remdesivir, found that COVID-19 patients, with symptom duration of ≤ 10 days, improved faster compared to that of the placebo group, but this finding was not statistically significant [ 166 ]. A study conducted in the USA, Europe, and Canada showed clinical improvement among severe COVID-19 hospitalized patients (36 of 53 patients; 68%) who were treated with compassionate use of remdesivir [ 167 ]. Another RCT that included 584 patients with moderate COVID-19 at 105 hospitals in the United States, Europe, and Asia found those who took a 5-day course of remdesivir compared with those randomized to standard care had a statistically better outcome [ 168 ]. However, the WHO Solidarity Trial [ 169 ], conducted in 30 countries, found that remdesivir (including hydroxychloroquine, lopinavir/ritonavir, and interferon) had little or no effect on overall mortality, ventilation need, and duration of hospital stay.
Although the preliminary findings of the ACTT-1 study supported the use of remdesivir, the researcher recommended that remdesivir or any other antiviral drug alone is not effective, as the mortality rate is higher with the use of remdesivir. Another randomized, controlled, open-label trial with lopinavir/ritonavir treatment demonstrated no benefit compared to standard care [ 170 ]. Similarly, the UK RECOVERY (Randomized Evaluation of COVID-19 therapy) reported no benefit of lopinavir/ritonavir on survival, the clinical course, or the length of hospital stay [ 171 ]. After interim analysis of the trial results, the WHO SOLIDARITY and UK RECOVERY trials discontinued the lopinavir/ritonavir arms as the trials produced little or no reduction in the mortality of hospitalized COVID-19 patients in comparison to the standard of care [ 172 , 173 ]. Remdesivir received conditional marketing authorization by the European Commission on 3 July 2020, to treat COVID-19 patients [ 174 ]. Several anti-flu drugs, such as oseltamivir [ 175 ] and arbidol [ 176 ], have been used to treat COVID-19 patients and demonstrated a certain efficacy. Although the WHO recommended against the use of remdesivir in COVID-19 patients [ 177 ], the U.S. Food and Drug Administration (FDA) approved remdesivir on 22 October 2020, for the treatment of COVID-19 patients requiring hospitalization [ 178 ]. Remdesivir (Veklury) was the first drug approved by the FDA and indicated ‘for the treatment of COVID-19 disease in hospitalized adults and children aged 12 years and older who weigh at least 40 kg’ [ 178 ].
8.3.2. Corticosteroids
Corticosteroids have received considerable attention for the treatment of COVID-19 [ 179 , 180 ] and were found to be beneficial in several COVID-19-related conditions such as sepsis, pneumonia, and ARDS [ 181–183 ]. The RECOVERY trial found that dexamethasone reduced mortality by one-third in critically ill COVID-19 patients [ 184 , 185 ]. The medication was most helpful for patients on a ventilator or those who needed extra oxygen, but no benefit was noted for those with less severe symptoms. However, other studies reported conflicting results, with some showing benefits [ 186–188 ], while others demonstrated potential harm [ 189 , 190 ]. A meta-analysis of 15 studies [ 191 ] identified an increased risk of mortality and multi-organ dysfunction, no mortality benefit, and possibly an increased risk of death with the use of corticosteroids among COVID-19, SARS, and MERS patients. A recent WHO report suggested that systemic corticosteroids likely reduced 28-day mortality in patients with critical COVID-19 but may have increased the risk of death in non-severe patients [ 174 ]. The report recommended the use of systemic corticosteroid therapy for 7 to 10 days in patients with severe and critical COVID-19 and no corticosteroid treatment for non-severe patients in whom it may cause harm.
8.3.3. Antiviral/immunomodulatory drugs
Chloroquine and hydroxychloroquine are usually used as immunomodulatory therapies. Both drugs are approved by the FDA for the treatment or prevention of malaria. Recently, the FDA has approved the use of chloroquine and hydroxychloroquine to treat COVID-19 patients ‘only in hospitalized patients with COVID-19 when clinical trials are not available, or participation is not feasible, through an Emergency Use Authorization (EUA)’ [ 192 ]. According to ClinicalTrials.gov, 212 hydroxychloroquine trials (179 randomized) and 38 chloroquine trials (31 randomized trials) were registered until 3 September 2020 [ 193 ].
However, the outcomes of treatment with chloroquine (500 mg every 12 h) and hydroxychloroquine are not encouraging. The RECOVERY trial found that hydroxychloroquine did not reduce 28-day mortality when compared to the usual standard of care. In addition, patients who received hydroxychloroquine had a longer median hospital stay and increased risk of progressing to invasive mechanical ventilation or death than those who received the standard of care [ 194 ]. In a multicenter, randomized, open-label, three-group, controlled trial involving hospitalized patients in Brazil, no positive outcomes were reported with hydroxychloroquine alone or with hydroxychloroquine plus azithromycin among hospitalized patients with mild to moderate COVID-19 [ 195 ]. The occurrence of an adverse event (e.g. elevation of liver enzyme levels, and prolongation of the QTc interval) was more frequent among patients who received hydroxychloroquine or hydroxychloroquine plus azithromycin than among those who did not receive either agent [ 195 ]. Another open-label, randomized clinical trial at 57 centers in Brazil involving hospitalized patients with severe COVID-19 also failed to show the effectiveness of hydroxychloroquine plus azithromycin over hydroxychloroquine alone [ 196 ]. Large retrospective observational studies in hospitalized patients suffering from COVID-19 also showed no evidence of benefit for hydroxychloroquine with or without azithromycin [ 197 , 198 ]. However, a large, multicenter, retrospective, observational study in the USA reported that treatment with hydroxychloroquine alone and in combination with azithromycin reduced COVID-19 associated mortality [ 199 ]. Several randomized trials conducted among non-hospitalized patients with COVID-19 failed to demonstrate a clinical benefit of hydroxychloroquine treatment [ 200 , 201 ].
The COVID-19 Treatment Guidelines of National Institutes of Health, USA recommends against the use of high-dose chloroquine to treat COVID-19 due to severe toxicities, such as higher rates of mortality and QTc prolongation [ 202 , 203 ]. It has been warned that the combination of hydroxychloroquine and azithromycin should be used with caution as the combination is associated with QTc prolongation in patients with COVID-19 [ 204 ].
8.3.4. Immune-based therapy
The agents that modulate the immune response are used for the management of moderate to critical COVID-19, including human blood-derived products and immunomodulatory therapies. Human blood-derived products are collected from patients who have recovered from COVID-19 infection (e.g. convalescent plasma and immunoglobulin products) [ 205 , 206 ]. Other agents approved to treat other immune and/or inflammatory syndromes are also considered to treat COVID-19 patients, including corticosteroids (e.g. glucocorticoids) [ 207 ], interleukin inhibitors [ 208 , 209 ], interferons [ 210 ], and kinase inhibitors [ 211 ].
It has been suggested that convalescent plasma may help suppress the virus and modify the inflammatory response [ 205 ]. At present, there is limited evidence from clinical trials to evaluate the efficacy and safety of convalescent plasma for the treatment of COVID-19 [ 212 ]. A retrospective evaluation conducted by the FDA and the Mayo Clinic (USA) in >70,000 patients who received COVID-19 convalescent plasma demonstrated that plasma with high antibody titers may be more effective than low-titer plasma in non-intubated patients [ 213 , 214 ]. The FDA also evaluated 20,000 hospitalized patients with COVID-19 convalescent plasma and reported that transfusion is safe in patients with COVID-19 and found low overall rates of serious adverse events (SAEs) [ 215 ]. It is important to note that the FDA approved EUA on 23 August 2020, to use convalescent plasma in hospitalized patients with COVID-19 [ 216 ].
Interferon β was found to be effective against coronaviruses [ 217 ]. The WHO Solidarity Trial [ 169 ] found that interferon had little or no effect on overall mortality, ventilation need, and duration of hospital stay. However, a randomized, double-blind, placebo-controlled phase 2 trial conducted in the UK [ 218 ] demonstrated that hospitalized patients infected with SARS-CoV-2 received inhaled nebulized interferon β-1a had significantly greater odds of clinical improvement and rapid recovery on the WHO ordinal scale for clinical improvement [ 219 ]. Further studies should be conducted to evaluate the effectiveness of high-risk COVID-19 populations such as elderly, comorbid, or immunosuppressed patients [ 169 , 218 ].
8.3.5. Adjunctive therapy
Adjunctive therapies are used in patients with COVID-19, and some clinical trials are ongoing to identify the effects of these agents [ 202 ]. It was observed that COVID-19 patients were associated with a prothrombotic state and had a higher incidence of venous thromboembolism [ 220 , 221 ]. A French prospective multicenter study among ICU patients (n = 150) demonstrated that 16.7% of patients with ARDS secondary to COVID-19 developed life-threatening thrombotic complications despite prophylactic anticoagulation [ 221 ]. Another study conducted in the Netherlands found a 31% incidence of thrombotic complications in critically ill ICU patients with COVID-19 (n = 184) [ 222 ]. Therefore, patients with COVID-19 admitted to the ICU should receive pharmacological thrombosis prophylaxis [ 222 ].
Vitamin and mineral supplements are typically used to treat respiratory viral infections. Several studies have examined the effectiveness of vitamin and mineral supplements for the treatment and prevention of SARS-CoV-2 infection. High doses of vitamin C are recommended for the treatment of sepsis [ 223 ] and ARDS in patients with serious COVID-19. Several recent studies have examined the impact of vitamin D on COVID-19. One study of 489 people found that those who had a deficient vitamin D status were 1.77 times more likely to be infected with the virus than people with normal vitamin D status [ 224 ]. Despite the lack of evidence of whether vitamin D treatment may decrease the incidence of COVID-19, the use of vitamin D treatment is advocated due to its low risk and low cost [ 225 , 226 ]. Some clinical trials are ongoing with zinc supplementation alone or in combination with hydroxychloroquine for the prevention and treatment of COVID-19 [ 227–230 ]. A single-institution retrospective study in the USA showed ‘a lack of a causal association between zinc and survival in hospitalized patients with COVID-19’ [ 231 ].
8.4. A vaccine
Scientists are conducting research on the development of COVID-19 vaccines. At present, there are >100 COVID-19 vaccine candidates under development, some of which are in the human trial phase [ 138 ]. The WHO is working through the Access to COVID-19 Tools (ACT) accelerator to speed up the pandemic response and distribute vaccines via the COVID-19 Vaccines Global Access (COVAX) [led by WHO, Global Alliance for Vaccines and Immunization (GAVI) and Coalition for Epidemic Preparedness Innovations (CEPI)] to facilitate equitable access and distribution [ 138 ]. The WHO announced the launch of the WHO COVID-19 Solidarity vaccine trial on 28 May 2020, which is an international, randomized controlled phase III trial of different vaccine candidates [ 232 ]. It is one of the largest trials that enrolled almost 280,000 patients from 470 hospital sites in over 34 countries [ 232 , 233 ]. The trial aims to examine the efficacy of multiple vaccines (within a short period of vaccine introduction into the study), so that weakly effective vaccines are not deployed to treat patients with COVID-19 [ 232–235 ].
Until October 2020, there were 42 COVID-19 candidate vaccines in the clinical evaluation, of which 10 were in phase 3 trials ( Table 4 ) [ 232 , 236 ]. There are 151 candidate vaccines for preclinical evaluation [ 232 ]. So far, four vaccines have been reported to be effective for the prevention of COVID-19: Pfizer/BioNtech, Moderna, Oxford, and Sputnik V vaccines. The details of these vaccines are presented in Table 4 [ 232 , 236 ] and Table 5 [ 139 , 141 , 237 , 238 ]. The first two vaccines received emergency approval for use in the prevention of COVID-19 [ 139–141 ]. Whether these vaccines are effective against new strains of SARS-CoV-2, which were recently identified in the UK and other countries, needs further investigation.
COVID-19 vaccine candidates in phase III trials [ 231 , 235 ]
| ** | |||
|---|---|---|---|
| Sinovac | Inactivated virus | S | Brazil |
| Wuhan Institute of Biological Products/Sinopharm | Inactivated virus | S | United Arab Emirates |
| Beijing Institute of Biological Products/Sinopharm | Inactivated virus | S | China |
| University of Oxford/AstraZeneca | Viral vector* | S | USA |
| CanSino Biological Inc./Beijing Institute of Biotechnology | Viral vector* | S | Pakistan |
| Gamaleya Research Institute | Viral vector | S | Russia |
| Janssen Pharmaceutical Companies | Viral vector | S | USA, Brazil, Colombia, Peru, Mexico, Philippines, South Africa |
| Novavax | Protein subunit | S | UK |
| Moderna/NIAID | RNA | S | USA |
| BioNTech/Fosun Pharma/Pfizer | RNA | S | USA, Argentina, Brazil |
* Single dose schedule. ** S – Spike protein
Vaccines found to be effective in preventing COVID-19 [ 139 , 141 , 237 , 238 ]
| * | |||||
|---|---|---|---|---|---|
| Viral vector (genetically modified virus) | 2 | IM | 62–90% | Regular fridge temperature | |
| RNA | 2 | IM | 95% | −70°C | |
| RNA (Part of virus genetic code) | 2 | IM | 95% | −20°C up to 6 months | |
| Gamaleya ( | Viral vector | 2 | IM | 92% | Regular fridge temperature |
*Intra-muscular
8.4.1. Pfizer/biontech vaccine
On 9 November 2020, Pfizer and its German partner BioNTech announced that their experimental vaccine was found to be more than 90% effective in preventing COVID-19 in participants without evidence of prior SARS-CoV-2 infection, based on initial data from Phase 3 trials [ 239 ]. According to Pfizer, the vaccine prevented COVID-19 symptoms in 90% of 94 patients who received the vaccine compared to the placebo. As of 8 November 2020, a total of 38,955 participants had received a second dose of the vaccine, of which 42% of global participants and 30% of U.S. participants had diverse racial and ethnic backgrounds [ 239 ]. Approximately 21% of the participants had at least one underlying comorbidity, that is, obesity, diabetes, or pulmonary disease [ 240 ]. On 16 November 2020, Pfizer released updated information concerning the observed efficacy of its vaccine in adults over 65 years of age, which was more than 94% [ 241 ]. On 11 December 2020, the FDA authorized the Pfizer/BioNTech vaccine for emergency use for individuals aged 16 years and older in the USA. This is the first COVID-19 vaccine approved by the FDA [ 139 ]. The European Medicines Agency (EMA) has also approved the Pfizer-BioNTech vaccine as the first COVID-19 vaccine to be used in EU countries [ 140 ].
The Pfizer/BioNTech vaccine is a messenger RNA (mRNA) vaccine, also known as BNT162b2, based on the SARS-CoV-2 S glycoprotein antigen and formulated in lipid nanoparticles (LNPs) [ 240 ]. It is a highly purified single-stranded, 5ʹ-capped mRNA produced by cell-free in vitro transcription from the corresponding DNA templates [ 242 ]. Its mechanism of action consists of nucleoside-modified mRNA (modRNA) encoding the viral S glycoprotein of SARS-CoV-2, which is formulated in lipid particles. This allows the delivery of RNA into host immune cells to enable the expression of the SARS-CoV-2 S antigen.
8.4.2. Moderna vaccine
On 16 November 2020, Moderna, Inc., a US pharmaceutical company, announced that its vaccine was 94.5% effective (Phase 3 COVE study) at preventing COVID-19 related illness, including severe cases, and is generally well tolerated [ 243 ]. An interim analysis of 95 cases (90 COVID-19 in the placebo group versus 5 cases in the mRNA-1273 group) demonstrated ‘a point estimate of vaccine efficacy of 94.5% (p < 0.0001)’ [ 243 ]. The Coronavirus Efficacy and Safety (COVE) trial, a randomized and placebo-controlled study, recruited 30,000 participants in the USA, aged 18 and above [ 243 ]. Unlike the Pfizer vaccine, it can be stored at standard refrigerator temperatures, which are available in doctors’ offices, pharmacies, and hospitals [ 244 ]. On 18 December 2020, the FDA issued an EUA for the Moderna vaccine for use in individuals 18 years of age and older in the USA [ 141 ].
The Moderna vaccine also used a similar technology to Pfizer/BioNTech. The active ingredient of the Moderna vaccine is a synthetic mRNA encoding the pre-fusion stabilized S glycoprotein of SARS-CoV-2. Both vaccines differ in their composition of LNP that encase the RNA; additionally, the RNA in both vaccines encodes a slightly modified form of the SARS-CoV-2 S protein [ 245 ]. Moderna’s formulation allows the vaccine to be stored at a higher temperature than Pfizer’s, which must be kept at −70°C, much colder than a normal freezer. Moderna’s vaccine can be stored in a − 20°C freezer for 6 months, and in a refrigerator (at approximately 4°C) for 30 days [ 141 , 237 ].
8.4.3. Oxford vaccine
Another vaccine developed by the University of Oxford, UK, and another pharmaceutical giant AstraZeneca was found highly effective – two full doses gave 62% protection (n = 8,895), a half dose followed by a full dose 90% (n = 2,741). Overall, the trial showed 70% protection (n = 11,636) [ 246 ]. The trial participants (n = 23,000) were from the UK and Brazil. The vaccine is cheaper than Pfizer and Moderna and does not require an ultra-cold storage and transport system [ 247 ]. As the vaccine was found to be more effective in trial participants who received a lower dose, AstraZeneca is now planning to run a new global trial [ 247 ].
Unlike the mRNA vaccines of Pfizer-BioNTech and Moderna, this vaccine uses double-stranded DNA. The mechanism of the vaccine is based on its effect on the S protein of SARS-CoV-2. The Oxford-AstraZeneca team used a modified version of the cold-causing chimpanzee adenovirus, known as ChAdOx1. Adenovirus derived from chimpanzee with E1 and E3 deletions encoding full-length S protein with a tissue plasminogen activator signal peptide [ 248 ]. With the use of genetic engineering methods, a portion of the DNA that is used for viral replication was deleted, so the adenovirus can no longer replicate and cause infection in the human body [ 249 ].
8.4.4. Sputnik V vaccine
The Russian vaccine Sputnik V was developed by the Gamaleya Research Institute in coordination with the Russian Defense Ministry. It was administered to 18,794 volunteers who received both the first and second doses of the vaccine or placebo. It showed very high efficacy; higher than 95% [ 238 ]. It is an adenovirus vector-based vaccine that uses a two-shot model with two different human adenoviral vectors, Ad5 and Ad26, for each shot [ 250 ]. When the first vaccine containing the vector with the S protein of SARS-CoV-2 is introduced into the human body, it synthesizes the S protein and initiates an immune response. After 21 days, the booster dose of the vaccine, based on another adenovirus vector unknown to the host cell, is administered. The body reacts by generating a further immune response that provides longlasting immunity [ 251 ].
9. The ethics of epidemics: ethical and moral issues associated with COVID-19
When it comes to global pandemics such as COVID-19, there are numerous issues in medical ethics that must be addressed and adhered to in order to ameliorate the human condition [ 252 , 253 ]. One of the most important issues to consider is patient confidentiality [ 254 ]. While confidentiality must be maintained between physicians and patients during standard medical care, when it comes to the treatment of a patient diagnosed with COVID-19, an exception has to be made. Since COVID-19 is considered a reportable illness, the type of illness that poses a threat to another person, doctors must follow Tarasoff’s Law of duty to warn and protect [ 255 ]. In other words, physicians are required to report quarantine and follow-up contact tracing [ 256 ].
Another ethical issue, autonomy, must be considered when a patient is diagnosed with COVID-19. A legally competent adult patient (18 years of age and older) may exercise their autonomous right to refuse treatment [ 257 ]. In such a case, a physician’s duty is to notify the patient about the possible health outcomes of refusing the treatment. However, COVID-19 is considered to be a quarantinable disease; thus, physicians could detain infected individuals during the infectious period.
In addition, informed consent is not required in the case of an emergency, such as in the case of a life-saving procedure for a patient diagnosed with COVID-19. Another example of an exception to obtaining informed consent is when the COVID-19 patient waived his or her right receiving information related to COVID-19 [ 254 ]. If a physician has to treat a patient diagnosed with COVID-19 who is incapacitated because he or she is either psychotic, unconscious, suicidal/homicidal, or under the influence, obtaining informed consent is not necessary [ 257 ]. Furthermore, physicians can invoke therapeutic privilege if physicians agree that the COVID-19 patient is unable to make good decisions for himself or herself. In this case, beneficence trumps the adult patient’s autonomy; hence, informed consent is not required during treatment [ 255 ].
In the case of minor health care (persons younger than 18 years of age), legally competent adult caregivers give consent for treatment [ 256 ]. Thus, when it comes to treating a minor diagnosed with COVID-19, the same rule applies as in the case of an adult patient. While physicians must always obtain informed consent from legal guardians when treating a minor, lifesaving treatment is always an exception [ 234 ]. Hence, legally competent adult guardians cannot refuse the lifesaving treatment of COVID-19 minors. On the other hand, when it comes to legally competent emancipated minors who are diagnosed with COVID-19, the physician must apply Tarasoff’s Law of duty to warn and protect [ 256 ]. In other words, the physicians must report, and quarantine emancipated minors diagnosed with COVID-19 since they pose a threat to another person and community. Furthermore, physicians could override their autonomous rights to refuse therapy by invoking therapeutic privileges, just in the case of adult patients [ 257 ].
10. Conclusion
The COVID-19 pandemic is the greatest global public health crisis since the pandemic influenza outbreak of 1918. Since its origin in Wuhan, the COVID-19 pandemic has now spread around the world, causing significant morbidity and mortality. Direct person-to-person respiratory transmission has rapidly amplified the spread of the virus, making it difficult to contain its spread within the community. Moreover, some patients are completely asymptomatic with a mild influenza-like illness and a positive swab test, and some present with serious symptoms that require immediate hospitalization. Currently, there is no effective antibody test available, and an effective, rapid, and sensitive serological test for COVID-19 is urgently needed for rapid diagnosis. Moreover, there is no effective approved therapy for COVID-19. Personal hygiene is fundamental for preventing transmission. Current treatment and management are mainly supportive of oxygen therapy, antivirals, steroids, hydroxychloroquine, immunomodulators, and plasma exchange therapy. We need to keep a close eye on human clinical trials for optimistic news on vaccine development.
10.1. Limitations
The information presented in this review paper must be considered in the context of potential limitations. There has been an overwhelming amount of information published since the outbreak of COVID-19, and most of the journal papers published, mainly in the early phase of pandemics, were not based on clinical/scientific research. The evidence-based information garnered for this review was obtained after careful review of currently published journal papers, reports, policy guidelines.
Another drawback of this paper is its narrative nature, which may limit the critical analysis of the information. A systematic review using appropriate protocols [ 258 ] of the current literature would help to draw reliable and accurate scientific information, improve the generalizability and consistency of findings, and increase the precision of the conclusion presented to formulate policy guidelines. However, the present review covers the most updated information on the anniversary of the COVID-19 pandemic, and such documentation is necessary for keeping readers, researchers, scientists, and policymakers appraised of the current status of the pandemic [ 259 ].
11. Expert Opinion
The COVID-19 pandemic has created a public health crisis, taking an enormous toll on humanity, disrupting lives and livelihoods [ 4 , 77 , 259 ]. The scale and severity of COVID-19 is unprecedented, and millions of people have been infected with large numbers of morbidities and mortalities [ 4 , 57 ]. Genetic sequencing suggests that the virus belongs to the family Coronaviridae and genus Betacoronavirus, which is closely linked to the SARS virus [ 27–30 ]. Epidemiological and virologic studies have reported that COVID-19 usually transmits from person to person through several routes, mainly via respiratory droplets [ 260–263 ]. Evidence of virological assessment of transmission of infection from people with presymptomatic stage is limited due to the lower number of samples collected [ 264 , 265 ]. Some infected persons can be contagious during the presymptomatic phase, from to 1–3 days before symptom onset [ 266 , 267 ]. For individuals, transmission risk is found to be greatest on the day of symptom onset in symptomatic patients [ 264 , 265 , 268 , 269 ]. Ferretti et al. analyzed five datasets and demonstrated that approximately 10% of transmissions may occur two days before the manifestation of symptoms [ 270 ]. Another review identified that 31% of infected individuals remain asymptomatic [ 271 ].
While most people with COVID-19 show only mild (40%) or moderate (40%) symptoms, approximately 15% of patients exhibit severe symptoms (requiring oxygen therapy), and 5% develop critical disease with complications (e.g. respiratory failure, ARDS, sepsis, septic shock) [ 135 ]. The WHO reported that the crude mortality ratio (the number of reported deaths divided by the reported cases) is 3–4%; however, the true mortality of COVID-19 will take some time to determine [ 271 ]. Elderly people, smokers, and patients with comorbid diseases (such as diabetes, hypertension, cardiac disease, chronic lung disease, and cancer) have an increased risk of severe disease and death [ 272 , 273 ].
The host response to SARS-CoV-2 is a key factor in the presentation of disease severity; however, variations in viral strain phenotypes, specifically those associated with the glycoprotein components of the virus, have contributed to the efficient transmission of the virus during the current pandemic [ 274 ].
Although the sequence diversity of SARS-CoV-2 is low, its global spread has resulted in several thousand viral variants due to mutations in the native strain over time [ 259 , 275 ]. The most notable of these, as first documented by Korber et al . [ 277 ], is a viral variant with an amino acid substitution in the S glycoprotein spike. The mutation, which causes a substitution of the amino acid aspartate (D-biochemical symbol), at the 614 th amino acid position of the spike protein with glycine (G), has overtaken the native SARS-CoV-2 virus as the most prevalent infective strain [ 277 ]. This variant, termed D614G, is associated with increased transmissibility and higher viral loads in COVID-19 patients, has not been demonstrated to cause an increase in disease severity [ 276 , 278 ]. The substitution enhances viral replication within the respiratory tract of infected individuals and affects neutralization susceptibility [ 274 ].
Compared with other highly mutable viruses, such as HIV, SARS-CoV-2 has a low mutation rate; however, as pandemics progress, it is possible that antigenic drift events, which slowly accumulate mutations over time, can lead to increased fitness as well as immunological and drug resistance [ 279 ]. This is a key consideration for current and future vaccine development.
Case detection, contact tracing, surveillance, infection prevention and control, physical distancing, and clinical management are effective strategies used to contain COVID-19 cases [ 280 , 281 ]. Early detection and reporting can prove to be useful, and contact tracing is a widely used surveillance system to fight the ongoing epidemic of COVID-19 [ 282 ]. Contact tracing provides information that also helps to better understand the transmission and epidemiology of COVID-19 [ 158 , 281 ]. Moreover, some countries are now experiencing the ‘second’ or ‘third’ waves of coronaviruses [ 283 ]. Scientists have proposed using app-based contact tracing to keep the epidemic in control as an alternative [ 284 ]. A digital technological system, called ‘proximity tracking ’ is now a widely used surveillance system for COVID-19 [ 285 ].
Currently, PCR, the gold standard for detecting SARS-CoV-2, is used to detect the virus in specialized laboratories [ 2 , 125 , 127 , 286 ]. The test has high sensitivity and specificity for the detection of viral ribonucleic acid (RNA) [ 2 , 286 ]. The high volume of samples could lead to a shortage of reagents and may increase the turn-around time of the tests. Alternatively, rapid antigen tests provide multiple benefits, including ease of use, quick results (10 to 30 minutes), low cost, and can be performed both in the laboratory and at the point of patient care [ 280 ]. Although rapid antigen testing has a lower sensitivity, the WHO recommends the use of this test where PCR is unavailable or where reduced turnaround-time is clinically necessary [ 287 ]. Antigen tests are immunoassays that are used to determine if the person has an active disease [ 288 ], whereas a positive antibody test indicates that the patient is likely infected with COVID-19 at some time in the past [ 289 ]. Antibody tests can be conducted in laboratory settings (e.g. enzyme-linked immunosorbent assays, chemiluminescence immunoassays) or point of care (e.g. Abbott SARS-CoV-2 assay, Roche Elecsys assay) [ 290 ].
To date, no effective specific drug therapy or vaccine has been found to limit the spread of this pathogen. Infection prevention and control measures, supportive needs, and intensive care support are the main strategies for clinical management of COVID-19 infection [ 136 ]. General prevention and protection measures regarding the containment and management of the second or third waves are necessary to minimize the risk of infection. There is some promising news regarding COVID-19 vaccines. Several phase 3 clinical trials are in progress or are being planned in some countries. As of 24 November 2020, four vaccines were reported to be 62–95% effective ( Table 5 ) [ 139 , 141 , 237 , 238 ]. These promising results have fueled optimism around the world, as we may be a step closer to defeating this deadly virus. Certainly, there is light at the end of the tunnel. Equitable access and effective distribution of these vaccines in all countries will save millions of lives.
ABBREVIATIONS
| COVID-19 - | Coronavirus disease 2019 |
| SARS-CoV-2 - | Severe acute respiratory syndrome coronavirus-2 |
| WHO - | World Health Organization |
| CVD - | Cardiovascular diseases |
| UTI - | Upper respiratory tract infections |
| HCoV - | Human coronavirus |
| SARS - | Severe acute respiratory syndrome coronavirus |
| MERS - | Middle East Respiratory Syndrome |
| ACE2 - | Angiotensin-converting enzyme 2 |
| RBD - | Receptor-binding domain |
| NTD - | N-terminal domain |
| R - | Reproduction number |
| ARDS - | Acute respiratory distress syndrome |
| PRRs - | Pattern recognition receptors |
| TLRs - | Toll-Like Receptors |
| CRP - | C-Reactive Protein |
| MODS - | Multiple organ dysfunction syndrome |
| CDCs - | Centres for Disease Control and Prevention |
| PCR - | Polymerase Chain Reaction |
| LAMP - | Loop-mediated isothermal amplification |
| EAU - | Emergency Use Authorization |
| ELISA - | Enzyme-Linked Immunosorbent Assay |
Funding Statement
This paper was not funded.
Article highlights
• The COVID-19 pandemic has created a public health crisis, infected millions of people, and caused a large number of morbidities and mortalities.
• COVID-19 transmits from person to person through several routes, mainly via respiratory droplets, which makes it difficult to contain its spread into the community.
• Currently, there is no effective antibody test available, and an effective, rapid, and sensitive serological test for COVID-19 is urgently needed for rapid diagnosis.
• In the absence of any clinically proven treatment options, the current clinical management of COVID-19 includes symptom management, infection prevention and control measures, optimized supportive care, and intensive care support for severe or critical illness, and general prevention and protection measures regarding the containment and management of the second or third waves are necessary to minimize the risk of infection.
• There is some promising news regarding COVID-19 vaccines as large-scale (Phase 3) clinical trials are in progress. As of 24 November 2020, four vaccines were reported to be 62–95% effective. Equitable access and effective distribution of these vaccines in all countries will save millions of lives.
Reviewer disclosures
Peer reviewers on this manuscript have no relevant financial or other relationships to disclose.
Declaration of interest
The authors have no relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript. This includes employment, consultancies, honoraria, stock ownership or options, expert testimony, grants or patents received or pending, or royalties.
Papers of special note have been highlighted as either of interest (•) or of considerable interest (••) to readers.
Newly Launched - AI Presentation Maker
Researched by Consultants from Top-Tier Management Companies

AI PPT Maker
Powerpoint Templates
Icon Bundle
Kpi Dashboard
Professional
Business Plans
Swot Analysis
Gantt Chart
Business Proposal
Marketing Plan
Project Management
Business Case
Business Model
Cyber Security
Business PPT
Digital Marketing
Digital Transformation
Human Resources
Product Management
Artificial Intelligence
Company Profile
Acknowledgement PPT
PPT Presentation
Reports Brochures
One Page Pitch
Interview PPT
All Categories
Top 25 Virus PowerPoint Templates To Beat the Invisible Threat

Gunjan Gupta
Viruses are curious little creatures. These tiny tagalongs aren’t exactly living per se, but they are not inanimate either. They’ve replicated themselves into multiple forms both fatal and non-fatal in billions of years and continue to do so even today. These beings have a long trail of history attached to their evolution and perhaps even had a hand or a squishy-squashy protein coating- in helping them mutate and become ever so dangerous. So, much so that they have become a threat to human existence.
In the recent turn of events, where the global pandemic Corona virus popularly known as Covid-19 is on a rise bringing along a chain of untimely deaths and mourns throughout the world, it is necessary to have a much closer look at these tiny microbial compounds.
If you are also curious to know about these tiny, yet not-so-tiny beings, their origination, evolution, health effects, and more, well tag along.
The evolutionary history of viruses is a fascinating, yet debatable topic for virologists and cell biologists, because of their diverse nature. As such it is a constant struggle to classify these entities and relate their existence to the human tree of life. They may be genetic elements or previously living organisms evolving as parasites. Constant research is going on these topics.
So, to support the vision of virologists and help them with their never-ending search, SlideTeam professionals have designed a few virus PowerPoint templates that will resolve some of the basic queries about these “microbes”. Each of these templates are apt to unfold new facts about viruses, and their existence, endangering the lives of the only living species in the world- us HUMANS!
Virus Templates to Download and Use
We are well aware that viruses are diverse in nature and so is their biological composition. Some have RNA genomes while some are made up of DNA genomes. This template can be used to explain the structure as well as the replication strategies of these microbes. It can also be used to explain the dizzying array of functions and health effects of viruses on human health as well as the research conducted on it. Some interesting facts about coronavirus and its genetic mutation can be unfolded in this section and presented to get a better understanding of the origin of this virus. These may include the 10-time mutation study of the virus, posing the biggest threat to humanity. Let alone, various other studies and scientific evidence can be presented by healthcare professionals by utilizing this interesting layout that can be customized.

Template 2
Science is the be-all and end-all of our findings. With the outbreak of the world deadliest virus ‘Coronavirus’, creating havoc in the lives of billions of people, this is a classic template that can be used by researchers and scientists to present their interesting studies. The biological composition of various viruses and their origination can be outlined and presented. This template can also be used by healthcare professionals to explain some of the symptoms and ill effects of various viruses. Not all the viruses are deadliest and so information about its various types can be elaborated on in this template. It also comes in handy for biological students to present their research findings after careful analysis of viruses, thus helping them impress their teachers.

Download Virus Science PowerPoint Templates
The Internet is flooded with false information about coronavirus and its outbreak. This template can be used by healthcare professionals to create awareness and stop the spreading of false rumors. It can also be used to present the recent worldwide stats of the infection as well as some safety measures that can be adopted by the masses to keep the spread in check. The science behind various viruses can be outlined in this template, to increase the overall understanding of the viewers. Nonetheless, it is an apt design for biomedical students to present their results, findings, and reviews on various viruses to advise medical practitioners.

Download Virus Science PowerPoint Templates And PowerPoint Background
Influenza is a major public health concern as it infects nearly 5-10% of the global population annually. Hence, there is a need to understand the molecular structure of this virus to reduce its strain on human health. Our professionally designed template can help you give a brief about how the virus multiples, infects, and how to combat infection, making it extremely useful for virologists and cell biologists. Additionally, it can be used to present researchers on few vaccines formulated to reduce the risk of influenza as well as the treatment prescribed. This design is fully adaptive to your needs so feel free to make any changes as you like depending upon your field of interest.

Download Structure Of The Influenza Virion Medical Images For PowerPoint
This template is ideal for a biology class to show and share important information on various viruses, their origin, infections caused by them, and others. The theme is formal and hence can be used to demonstrate any topic in science class and submit an assignment as well. Doctors, nurses and paramedical staff can also utilize this template to present the information on vaccine reactions which may happen to patients, thus creating awareness. A lot of studies conducted on viruses is also the topic this slide caters to. Additionally, such medical templates are apt for drug health lessons in ordinary schools, colleges, and medical universities. All of these characteristics make this a universally efficient template. Hence, any individual can download it to present his topic in a comprehensive manner.

Download Virus Microorganism Geographical PowerPoint Templates And PowerPoint Backgrounds
The outbreak of coronavirus throughout the world has become one of the most notorious events of the decade, if not the current century. We can say that we have welcomed a new decade with a catastrophe of death and mourning. Since, this pandemic is nothing short of deadly, taking away lakhs of lives, every bit of information helps a lot. This virus danger template can help you create awareness on this global pandemic. It is a virus and microorganisms based theme template to put everything topic related to the context. The control and prevention of this disease can be the pillars of your talking points after utilizing this presentation theme. Also, there is a consistency in the theme of this template and includes font size that can be easily viewed from a distance. Thus, making it an efficient design that can help you present great presentations on science, biology, virology, and other related fields.

Download Virus Danger Globe PowerPoint Templates And PowerPoint Backgrounds
This template is designed for HIV and Aids doctors, students, academicians, and other professionals looking for comprehensive designs to create presentations for their universities, hospitals, seminars, and several others. HIV and Aids is a very important topic for medical practitioners to get acquainted with. This template can help you showcase important information on Aids and the causes of its spread. This is also a great design for medics and professionals to convey their thoughts on the topic, thus helping them become better communicators.

HIV Virus Medical PowerPoint Templates And PowerPoint Backgrounds
The mechanism of HIV entry to the host cell can be explained with this pre-designed template. This layout focuses on the design needs of medical professionals to give a descriptive view of the topic. It is apt for doctors, nurses, paramedics, researchers, academicians, and others working in the field of HIV Aids and its transmission. This content-ready template enables you to deliver great presentations revolving around the topic of HIV with minimum effort and maximum effect. Its simple to download layout requires only a few clicks to get you ready for your upcoming medical presentations. Hence, convey your thoughts and ideas on HIV with this template, that is subjected to numerous alterations, depending upon the requirements of the user.

Download HIV Attachment To Target T Cell Medical Images For PowerPoint
Classification and explanation of the nomenclature of viruses is a breeze with this template. It is specifically designed for paramedical staff to discuss their views on Hepatitis B and how it affects the liver of the infected patient. Since almost 1 million people die every year because of chronic hepatitis B infection which is preventable and treatable this template comes in very handy to present the important details and facts. Also, new perspectives on the life cycle of hepatitis B can be thoroughly explained in medical universities with this thoughtfully designed template. Since this template is versatile and flexible it is easily adaptable to all your needs and requirements. All you have to do is click on the download button and use it extensively.

Download Hepatitis B Virus Medical Images For PowerPoint
Template 10
This template can be used to explain the microscopic view and nomenclature of the Papillomavirus or HPV infection. It can also be used to understand its preventive measures, causes, and treatment making it an ideal fit for doctors to give a deeper understanding of the topic. Hospitals and medical universities can utilize this template to teach and train students and staff members. This slide acts as a very good teaching tool as the viewers are able to get a thorough understanding of health care issues, proposed safety measures, reasons for infections, and similar other topics. Additionally, the pre-designed layout of this template showing the entire structure of the virus makes it extremely easy to deliver great medical presentations with minimum effort.

Download Human Papilloma Virus Medical Images For PowerPoint
Template 11
This template can be used by doctors, medical universities, researchers, virologists, cell biologists, and others to present a thorough understanding of the structure of HIV and how it leads to a non-treatable infection. It showcases two types of infection latent and active and how the synthesis happens. Since this template is so descriptive designed using apt color scheme, it is efficient enough to grab this immediate attention of the viewer. You can either use this template in conjunction with your presentation or as a single entity as it is complete in itself.

Download Latent And Active Infection Of T Cell By HIV Medical Images For PowerPoint
Template 12
Adenovirus is ubiquitous viruses that cause various pediatric illnesses including common cold. They can also cause gastrointestinal, ophthalmologic, neurologic, and genitourinary symptoms. This template has especially been designed for paramedics and cell biologists to explain the basic structure of adenovirus and its prevention. While illness caused due to viruses have become quite common, having a thorough understanding of the illness is important. This template can be used in a discussion session needing to study the causes of viral growth, its production, and other aspects. Due to its attractive layout, it never fails to impress any audience.

Download Adenovirus Morphology Medical Images For PowerPoint
Template 13
This is yet another adenovirus template that is suitable to discuss the structure and movement of viruses in the human body. You can also explain the causes of its growth, infection and expansion with this template that is fully adaptable to one’s needs and requirements. Biological experiments and genetic engineering projects can also be presented using this template that is fully responsive.

Download Adenovirus Medical Images For PowerPoint
Template 14
Download this Leukemia template for your upcoming medical presentations. This template is specifically designed keeping in mind the needs of medical practitioners, healthcare professionals, and pathologists. So, you can easily edit it with your own content as this template uses royalty-free images. Apart from this, the template design can be used as a background of your presentation on leukemia, symptoms, causes, types, diagnosis, and treatment. Therefore grab this multifunctional PPT now to convey your information on leukemia cancer impressively.

Download Leukemia Science PowerPoint Templates And PowerPoint Backgrounds
Template 15
Another family of virus Hepatitis A can be influentially explained with this pre-designed template. Doctors, nurses, and other medical staff members can use it to create presentations for universities, hospitals, and other places depending upon their needs and requirements. It is also a good explanation tool for conferences and medical counseling sessions conducted in hospitals. The causes, vaccine reactions, side effects of the vaccine, diagnosis, treatment, and other aspects can be discussed efficiently with this neatly-designed template. The pre-designed elements of this template make your presentation work quick and a lot more convenient.

Download Hepatitis A Virus Medical Images For PowerPoint
Template 16
Showcase important information related to Herpes Simplex Virus using this slide. Explain its causes, risk factors, symptoms, vaccine, diagnosis, treatment, and other important aspects. Since this slide caters to the biology of viruses it will act as a great teaching tool in schools, medical universities, hospitals, and other places. Apart from this, it can be used by any medical practitioner and cell biologists depending upon what their project commands.

Download Herpes Simplex Virus Medical Images For PowerPoint
Template 17
This pre-made template can be used to present amazing presentations on topics like virology, immunization, vaccination, and others. This template specifically targets Respiratory Syncytial Virus hence it can be used to give a thorough explanation of this virus, its types, symptoms, diagnosis and other aspects making it highly useful for the medical practitioners.

Download Respiratory Syncytial Virus Medical Images For PowerPoint
Template 18
This presentation design is perfect for presenting strategies on the stop of growth and spreading of infection in humans. You can outline the entire structure of Measles and the infection caused by it using this template. Doctors can also use this template to educate their patients about Measles and prescribe an effective treatment for it. Nonetheless, the layout of this slide is designed professionally hence it is an ideal fir for medical students as well.

Download Measles Virus Medical Images For PowerPoint
Template 19
Such templates are great for providing health lessons on various viruses, their causes, symptoms, and the like. They can be used by medical professionals, biology teachers, medical universities, and others as a contribution to preventing infections from spreading by creating awareness. Since this template is pre-designed it will function as a great tool to present medical presentations.

Download Arena Virus Particle Structure Medical Images For PowerPoint
Template 20
Give a descriptive presentation on the highly contagious and deadly disease Smallpox using this slide. Showcase the structure of variola virus and its infection that leads to smallpox in humans. You can also present the history and evolution of this virus along with its treatment. Use it in conjunction with your medical presentation to create an impact. The high-resolution images used in each of the slides act as great teaching tools for educators, academicians, paramedical staff, and others.

Download Smallpox Virus Medical Images For PowerPoint
Template 21
Many infectious diseases are becoming fatal among human beings because of the overgrowth of viruses. One such virus is the rubella virus causing red rashes throughout the body. This template aims at giving an overview of rubella, its causes, symptoms, and others. Thus, making it a highly resourceful design for your medical presentations.

Download Rubella Virus Medical Images For PowerPoint
Template 22
We are bombarded with new infections every year some which are deadly and some which are not. This presentation aims at targeting a similar virus and its infections known as the RSV virus causing respiratory issues in children. The descriptive layout of this template makes it apt for presenting various medical presentations that are impressive and convince enough. These templates are specially designed to ease the work of the user by providing ready-made design and aesthetics.

Download RSV Virus Medical Images For PowerPoint
Template 23
Mimivirus questions our understanding that viruses are small as this the largest and most complex virus ever found. This template can be used by presenters to explain the evolution, discovery, and analysis of mimivirus both from the scientific and medical perspectives. It showcases the complex structure of mimivirus and its characteristics which defy the conventional definitions of life. Using this template will help you explain the mimivirus more descriptively, thus making it a great template for your presentations.

Download Mimi Virus Particle Structure Medical Images For PowerPoint
Template 24
This template shows the entire lifecycle of HIV and its replication, thus making it an outstanding explanation tool. This template is fully functional for doctors, medical universities, teachers, academicians, etc. looking to present important information on HIV and its infection. It is also a great tool to create awareness about this particular virus among the masses so that necessary steps can be taken to keep its spread in check. Apart from this biological experiment lab and genetic engineering projects can also use this template in whichever way they prefer.

Download Replication Cycle Of Human Immunodeficiency Virus HIV Medical Images For PowerPoint
Template 25
While the world is battling with a pandemic Coronavirus and its mutated offsprings, we have designed this template to help you present some safety measures that will help fight with this virus, eating up the entire human race slowly and gradually. This template can be used to present information about the virus and safety measures to keep its spread in check like using sanitizers, wearing masks, observing social distancing, washing hands, not touching nose and face, etc. You can also use this template as a background for your presentations related to viruses.

Download Fight With Virus Medical PowerPoint Templates And PowerPoint Backgrounds
Contemplating the origin of life is a fascinating topic for both scientists and the general public such as you and me. Understanding the evolutionary history of viruses and their life span can surely shed some light on this topic, which is what these 25 virus templates aim for.
Although there is no clear explanation of the origin of these viruses, these have been present for millions of years among us humans. So, even a little bit of information can make a world of difference here.
Download these templates to spark a sense of hope in these difficult and dark times where our lives and deaths are dependent on a VIRUS called Coronavirus or COVID 19.
Related posts:
- 10 Best Medical Google Slides Templates For Improving Lives
- Increase The Cohesiveness Of Workplace With Our Top 20 Organizational Culture Presentation Templates for PowerPoint!!
- Top 35 Change Management Templates To Transform Your Business WorkFlow
- 9 Beautiful Color Palettes for Designing Powerful PowerPoint Slides
Liked this blog? Please recommend us

Top 40 Medical, Healthcare Templates for Doctors and Nurses

Top 25 Drugs and Medicines PowerPoint Templates trusted by Medical Professionals

Top 30 Medical PowerPoint Slides for the Soldiers Fighting Against CoronaVirus!

Battling Coronavirus: Dumbledore’s Wise Message Guides Us in These Dark Times

The Complete COVID-19 PPT Presentation: Outbreak & Impact
This form is protected by reCAPTCHA - the Google Privacy Policy and Terms of Service apply.

--> Digital revolution powerpoint presentation slides

--> Sales funnel results presentation layouts
--> 3d men joinning circular jigsaw puzzles ppt graphics icons

--> Business Strategic Planning Template For Organizations Powerpoint Presentation Slides

--> Future plan powerpoint template slide

--> Project Management Team Powerpoint Presentation Slides

--> Brand marketing powerpoint presentation slides

--> Launching a new service powerpoint presentation with slides go to market

--> Agenda powerpoint slide show

--> Four key metrics donut chart with percentage

--> Engineering and technology ppt inspiration example introduction continuous process improvement

--> Meet our team representing in circular format

Information
- Author Services
Initiatives
You are accessing a machine-readable page. In order to be human-readable, please install an RSS reader.
All articles published by MDPI are made immediately available worldwide under an open access license. No special permission is required to reuse all or part of the article published by MDPI, including figures and tables. For articles published under an open access Creative Common CC BY license, any part of the article may be reused without permission provided that the original article is clearly cited. For more information, please refer to https://www.mdpi.com/openaccess .
Feature papers represent the most advanced research with significant potential for high impact in the field. A Feature Paper should be a substantial original Article that involves several techniques or approaches, provides an outlook for future research directions and describes possible research applications.
Feature papers are submitted upon individual invitation or recommendation by the scientific editors and must receive positive feedback from the reviewers.
Editor’s Choice articles are based on recommendations by the scientific editors of MDPI journals from around the world. Editors select a small number of articles recently published in the journal that they believe will be particularly interesting to readers, or important in the respective research area. The aim is to provide a snapshot of some of the most exciting work published in the various research areas of the journal.
Original Submission Date Received: .
- Active Journals
- Find a Journal
- Proceedings Series
- For Authors
- For Reviewers
- For Editors
- For Librarians
- For Publishers
- For Societies
- For Conference Organizers
- Open Access Policy
- Institutional Open Access Program
- Special Issues Guidelines
- Editorial Process
- Research and Publication Ethics
- Article Processing Charges
- Testimonials
- Preprints.org
- SciProfiles
- Encyclopedia
Article Menu

- Subscribe SciFeed
- Recommended Articles
- Google Scholar
- on Google Scholar
- Table of Contents
Find support for a specific problem in the support section of our website.
Please let us know what you think of our products and services.
Visit our dedicated information section to learn more about MDPI.
JSmol Viewer
Host innate antiviral response to influenza a virus infection: from viral sensing to antagonism and escape.

1. Introduction
2. sensing of iav infection by host cells, 2.1. toll-like receptors (tlrs), 2.2. retinoic acid-inducible gene-like receptors (rlrs), 2.3. z-dna-binding protein 1 (zbp1), 2.4. nod-like receptors (nlrs), 3. downstream innate immune signaling against iav, 3.1. tlr-mediated downstream signaling, 3.2. rlr-mediated downstream signaling, 3.3. zbp1-mediated downstream signaling, 3.4. nlrp3-mediated downstream signaling, 3.5. irfs in downstream signaling, 3.6. nf-κb in downstream signaling, 3.7. interferons (ifns), 3.8. jak-stat pathway, 3.10. pro-inflammatory cytokines, chemokines, and growth factors, 4. antagonism of host innate immune response by iav, 4.1. antagonism of rig-i, 4.2. antagonism of irfs and nf-κb, 4.3. antagonism of nlrp3, 4.4. antagonism of jak-stat pathway, 4.5. antagonism of isgs, 5. iav’s escape from the host innate immune response, 5.1. escape from tlr- and rlr-mediated sensing, 5.2. escape from irf-mediated signaling, 5.3. escape from isg-mediated restriction, author contributions, acknowledgments, conflicts of interest.
- Riera Romo, M.; Perez-Martinez, D.; Castillo Ferrer, C. Innate immunity in vertebrates: An overview. Immunology 2016 , 148 , 125–139. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Beutler, B. Innate immunity: An overview. Mol. Immunol. 2004 , 40 , 845–859. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Iwama, R.E.; Moran, Y. Origins and diversification of animal innate immune responses against viral infections. Nat. Ecol. Evol. 2023 , 7 , 182–193. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Bergeron, H.C.; Hansen, M.R.; Tripp, R.A. Interferons-Implications in the Immune Response to Respiratory Viruses. Microorganisms 2023 , 11 , 2179. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Zhu, J.; Chiang, C.; Gack, M.U. Viral evasion of the interferon response at a glance. J. Cell Sci. 2023 , 136 , jcs260682. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Wei, L.; Wang, X.; Zhou, H. Interaction among inflammasome, PANoptosise, and innate immune cells in infection of influenza virus: Updated review. Immun. Inflamm. Dis. 2023 , 11 , e997. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Stroz, S.; Kosiorek, P.; Stasiak-Barmuta, A. The COVID-19 inflammation and high mortality mechanism trigger. Immunogenetics 2024 , 76 , 15–25. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- King, A.M.Q.; Adams, M.J.; Carstens, E.B.; Lefkowitz, E.J. (Eds.) Orthomyxoviridae. In Virus Taxonomy ; Elsevier: San Diego, CA, USA, 2012; pp. 749–761. [ Google Scholar ]
- Su, S.; Fu, X.; Li, G.; Kerlin, F.; Veit, M. Novel Influenza D virus: Epidemiology, pathology, evolution and biological characteristics. Virulence 2017 , 8 , 1580–1591. [ Google Scholar ] [ CrossRef ]
- Russell, C.A.; Fouchier, R.A.M.; Ghaswalla, P.; Park, Y.; Vicic, N.; Ananworanich, J.; Nachbagauer, R.; Rudin, D. Seasonal influenza vaccine performance and the potential benefits of mRNA vaccines. Hum. Vaccin. Immunother. 2024 , 20 , 2336357. [ Google Scholar ] [ CrossRef ]
- Long, J.S.; Mistry, B.; Haslam, S.M.; Barclay, W.S. Host and viral determinants of influenza A virus species specificity. Nat. Rev. Microbiol. 2019 , 17 , 67–81. [ Google Scholar ] [ CrossRef ]
- Kang, M.; Wang, L.-F.; Sun, B.-W.; Wan, W.-B.; Ji, X.; Baele, G.; Bi, Y.-H.; Suchard, M.A.; Lai, A.; Zhang, M.; et al. Zoonotic infections by avian influenza virus: Changing global epidemiology, investigation, and control. Lancet Infect. Dis. 2024 . Online . [ Google Scholar ] [ CrossRef ]
- Tang, C.Y.; Ramesh, A.; Wan, X.-F. Avian and swine influenza viruses. In Molecular Medical Microbiology ; Tang, Y.-W., Hindiyeh, M.Y., Liu, D., Sails, A., Spearman, P., Zhang, J.-R., Eds.; Academic Press: Cambridge, MA, USA, 2024; pp. 2375–2411. [ Google Scholar ]
- Dou, D.; Revol, R.; Ostbye, H.; Wang, H.; Daniels, R. Influenza A Virus Cell Entry, Replication, Virion Assembly and Movement. Front. Immunol. 2018 , 9 , 1581. [ Google Scholar ] [ CrossRef ]
- Li, H.C.; Yang, C.H.; Lo, S.Y. Strategies of Influenza A Virus to Ensure the Translation of Viral mRNAs. Pathogens 2022 , 11 , 1521. [ Google Scholar ] [ CrossRef ]
- Anderson, K.V.; Jurgens, G.; Nusslein-Volhard, C. Establishment of dorsal-ventral polarity in the Drosophila embryo: Genetic studies on the role of the Toll gene product. Cell 1985 , 42 , 779–789. [ Google Scholar ] [ CrossRef ]
- Chiang, C.; Beachy, P.A. Expression of a novel Toll-like gene spans the parasegment boundary and contributes to hedgehog function in the adult eye of Drosophila. Mech. Dev. 1994 , 47 , 225–239. [ Google Scholar ] [ CrossRef ]
- Lemaitre, B.; Nicolas, E.; Michaut, L.; Reichhart, J.M.; Hoffmann, J.A. The dorsoventral regulatory gene cassette spatzle/Toll/cactus controls the potent antifungal response in Drosophila adults. Cell 1996 , 86 , 973–983. [ Google Scholar ] [ CrossRef ]
- Medzhitov, R.; Preston-Hurlburt, P.; Janeway, C.A., Jr. A human homologue of the Drosophila Toll protein signals activation of adaptive immunity. Nature 1997 , 388 , 394–397. [ Google Scholar ] [ CrossRef ]
- Rock, F.L.; Hardiman, G.; Timans, J.C.; Kastelein, R.A.; Bazan, J.F. A family of human receptors structurally related to Drosophila Toll. Proc. Natl. Acad. Sci. USA 1998 , 95 , 588–593. [ Google Scholar ] [ CrossRef ]
- Hoshino, K.; Takeuchi, O.; Kawai, T.; Sanjo, H.; Ogawa, T.; Takeda, Y.; Takeda, K.; Akira, S. Cutting edge: Toll-like receptor 4 (TLR4)-deficient mice are hyporesponsive to lipopolysaccharide: Evidence for TLR4 as the Lps gene product. J. Immunol. 1999 , 162 , 3749–3752. [ Google Scholar ] [ CrossRef ]
- Poltorak, A.; He, X.; Smirnova, I.; Liu, M.Y.; Van Huffel, C.; Du, X.; Birdwell, D.; Alejos, E.; Silva, M.; Galanos, C.; et al. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: Mutations in Tlr4 gene. Science 1998 , 282 , 2085–2088. [ Google Scholar ] [ CrossRef ]
- Qureshi, S.T.; Lariviere, L.; Leveque, G.; Clermont, S.; Moore, K.J.; Gros, P.; Malo, D. Endotoxin-tolerant mice have mutations in Toll-like receptor 4 (Tlr4). J. Exp. Med. 1999 , 189 , 615–625. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Kirschning, C.J.; Wesche, H.; Merrill Ayres, T.; Rothe, M. Human toll-like receptor 2 confers responsiveness to bacterial lipopolysaccharide. J. Exp. Med. 1998 , 188 , 2091–2097. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Yang, R.B.; Mark, M.R.; Gray, A.; Huang, A.; Xie, M.H.; Zhang, M.; Goddard, A.; Wood, W.I.; Gurney, A.L.; Godowski, P.J. Toll-like receptor-2 mediates lipopolysaccharide-induced cellular signalling. Nature 1998 , 395 , 284–288. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014 , 5 , 461. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Botos, I.; Segal, D.M.; Davies, D.R. The structural biology of Toll-like receptors. Structure 2011 , 19 , 447–459. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Lester, S.N.; Li, K. Toll-like receptors in antiviral innate immunity. J. Mol. Biol. 2014 , 426 , 1246–1264. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Guillot, L.; Le Goffic, R.; Bloch, S.; Escriou, N.; Akira, S.; Chignard, M.; Si-Tahar, M. Involvement of toll-like receptor 3 in the immune response of lung epithelial cells to double-stranded RNA and influenza A virus. J. Biol. Chem. 2005 , 280 , 5571–5580. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Le Goffic, R.; Balloy, V.; Lagranderie, M.; Alexopoulou, L.; Escriou, N.; Flavell, R.; Chignard, M.; Si-Tahar, M. Detrimental contribution of the Toll-like receptor (TLR)3 to influenza A virus-induced acute pneumonia. PLoS Pathog. 2006 , 2 , e53. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Diebold, S.S.; Kaisho, T.; Hemmi, H.; Akira, S.; Reis e Sousa, C. Innate antiviral responses by means of TLR7-mediated recognition of single-stranded RNA. Science 2004 , 303 , 1529–1531. [ Google Scholar ] [ CrossRef ]
- Lund, J.M.; Alexopoulou, L.; Sato, A.; Karow, M.; Adams, N.C.; Gale, N.W.; Iwasaki, A.; Flavell, R.A. Recognition of single-stranded RNA viruses by Toll-like receptor 7. Proc. Natl. Acad. Sci. USA 2004 , 101 , 5598–5603. [ Google Scholar ] [ CrossRef ]
- Wang, J.P.; Bowen, G.N.; Padden, C.; Cerny, A.; Finberg, R.W.; Newburger, P.E.; Kurt-Jones, E.A. Toll-like receptor-mediated activation of neutrophils by influenza A virus. Blood 2008 , 112 , 2028–2034. [ Google Scholar ] [ CrossRef ]
- Le Goffic, R.; Pothlichet, J.; Vitour, D.; Fujita, T.; Meurs, E.; Chignard, M.; Si-Tahar, M. Cutting Edge: Influenza A virus activates TLR3-dependent inflammatory and RIG-I-dependent antiviral responses in human lung epithelial cells. J. Immunol. 2007 , 178 , 3368–3372. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Tsai, S.Y.; Segovia, J.A.; Chang, T.H.; Morris, I.R.; Berton, M.T.; Tessier, P.A.; Tardif, M.R.; Cesaro, A.; Bose, S. DAMP molecule S100A9 acts as a molecular pattern to enhance inflammation during influenza A virus infection: Role of DDX21-TRIF-TLR4-MyD88 pathway. PLoS Pathog. 2014 , 10 , e1003848. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Imai, Y.; Kuba, K.; Neely, G.G.; Yaghubian-Malhami, R.; Perkmann, T.; van Loo, G.; Ermolaeva, M.; Veldhuizen, R.; Leung, Y.H.; Wang, H.; et al. Identification of oxidative stress and Toll-like receptor 4 signaling as a key pathway of acute lung injury. Cell 2008 , 133 , 235–249. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Lee, S.M.; Kok, K.H.; Jaume, M.; Cheung, T.K.; Yip, T.F.; Lai, J.C.; Guan, Y.; Webster, R.G.; Jin, D.Y.; Peiris, J.S. Toll-like receptor 10 is involved in induction of innate immune responses to influenza virus infection. Proc. Natl. Acad. Sci. USA 2014 , 111 , 3793–3798. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Rehwinkel, J.; Gack, M.U. RIG-I-like receptors: Their regulation and roles in RNA sensing. Nat. Rev. Immunol. 2020 , 20 , 537–551. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Sun, Y.W. RIG-I, a Human Homolog Gene of RNA Helicase, Is Induced by Retinoic Acid during the Differentiation of Acute Promyelocytic Luekemia Cell. Thesis, Shanghai Institute of Heamatology, Rui-Jin Hospital, Shanghai Second Medical University, Shanghai, China, 1997. [ Google Scholar ]
- Liu, T.X.; Zhang, J.W.; Tao, J.; Zhang, R.B.; Zhang, Q.H.; Zhao, C.J.; Tong, J.H.; Lanotte, M.; Waxman, S.; Chen, S.J.; et al. Gene expression networks underlying retinoic acid-induced differentiation of acute promyelocytic leukemia cells. Blood 2000 , 96 , 1496–1504. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Imaizumi, T.; Aratani, S.; Nakajima, T.; Carlson, M.; Matsumiya, T.; Tanji, K.; Ookawa, K.; Yoshida, H.; Tsuchida, S.; McIntyre, T.M.; et al. Retinoic acid-inducible gene-I is induced in endothelial cells by LPS and regulates expression of COX-2. Biochem. Biophys. Res. Commun. 2002 , 292 , 274–279. [ Google Scholar ] [ CrossRef ]
- Zhang, X.; Wang, C.; Schook, L.B.; Hawken, R.J.; Rutherford, M.S. An RNA helicase, RHIV -1, induced by porcine reproductive and respiratory syndrome virus (PRRSV) is mapped on porcine chromosome 10q13. Microb. Pathog. 2000 , 28 , 267–278. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Yoneyama, M.; Kikuchi, M.; Natsukawa, T.; Shinobu, N.; Imaizumi, T.; Miyagishi, M.; Taira, K.; Akira, S.; Fujita, T. The RNA helicase RIG-I has an essential function in double-stranded RNA-induced innate antiviral responses. Nat. Immunol. 2004 , 5 , 730–737. [ Google Scholar ] [ CrossRef ]
- Kato, H.; Sato, S.; Yoneyama, M.; Yamamoto, M.; Uematsu, S.; Matsui, K.; Tsujimura, T.; Takeda, K.; Fujita, T.; Takeuchi, O.; et al. Cell type-specific involvement of RIG-I in antiviral response. Immunity 2005 , 23 , 19–28. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Kato, H.; Takeuchi, O.; Sato, S.; Yoneyama, M.; Yamamoto, M.; Matsui, K.; Uematsu, S.; Jung, A.; Kawai, T.; Ishii, K.J.; et al. Differential roles of MDA5 and RIG-I helicases in the recognition of RNA viruses. Nature 2006 , 441 , 101–105. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Hornung, V.; Ellegast, J.; Kim, S.; Brzozka, K.; Jung, A.; Kato, H.; Poeck, H.; Akira, S.; Conzelmann, K.K.; Schlee, M.; et al. 5′-Triphosphate RNA is the ligand for RIG-I. Science 2006 , 314 , 994–997. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Pichlmair, A.; Schulz, O.; Tan, C.P.; Naslund, T.I.; Liljestrom, P.; Weber, F.; Reis e Sousa, C. RIG-I-mediated antiviral responses to single-stranded RNA bearing 5′-phosphates. Science 2006 , 314 , 997–1001. [ Google Scholar ] [ CrossRef ]
- Goubau, D.; Schlee, M.; Deddouche, S.; Pruijssers, A.J.; Zillinger, T.; Goldeck, M.; Schuberth, C.; Van der Veen, A.G.; Fujimura, T.; Rehwinkel, J.; et al. Antiviral immunity via RIG-I-mediated recognition of RNA bearing 5′-diphosphates. Nature 2014 , 514 , 372–375. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Schlee, M.; Roth, A.; Hornung, V.; Hagmann, C.A.; Wimmenauer, V.; Barchet, W.; Coch, C.; Janke, M.; Mihailovic, A.; Wardle, G.; et al. Recognition of 5′ triphosphate by RIG-I helicase requires short blunt double-stranded RNA as contained in panhandle of negative-strand virus. Immunity 2009 , 31 , 25–34. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Schmidt, A.; Schwerd, T.; Hamm, W.; Hellmuth, J.C.; Cui, S.; Wenzel, M.; Hoffmann, F.S.; Michallet, M.C.; Besch, R.; Hopfner, K.P.; et al. 5′-triphosphate RNA requires base-paired structures to activate antiviral signaling via RIG-I. Proc. Natl. Acad. Sci. USA 2009 , 106 , 12067–12072. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Kato, H.; Takeuchi, O.; Mikamo-Satoh, E.; Hirai, R.; Kawai, T.; Matsushita, K.; Hiiragi, A.; Dermody, T.S.; Fujita, T.; Akira, S. Length-dependent recognition of double-stranded ribonucleic acids by retinoic acid-inducible gene-I and melanoma differentiation-associated gene 5. J. Exp. Med. 2008 , 205 , 1601–1610. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Baum, A.; Sachidanandam, R.; Garcia-Sastre, A. Preference of RIG-I for short viral RNA molecules in infected cells revealed by next-generation sequencing. Proc. Natl. Acad. Sci. USA 2010 , 107 , 16303–16308. [ Google Scholar ] [ CrossRef ]
- Rehwinkel, J.; Tan, C.P.; Goubau, D.; Schulz, O.; Pichlmair, A.; Bier, K.; Robb, N.; Vreede, F.; Barclay, W.; Fodor, E.; et al. RIG-I detects viral genomic RNA during negative-strand RNA virus infection. Cell 2010 , 140 , 397–408. [ Google Scholar ] [ CrossRef ]
- Yin, X.; Riva, L.; Pu, Y.; Martin-Sancho, L.; Kanamune, J.; Yamamoto, Y.; Sakai, K.; Gotoh, S.; Miorin, L.; De Jesus, P.D.; et al. MDA5 Governs the Innate Immune Response to SARS-CoV-2 in Lung Epithelial Cells. Cell Rep. 2021 , 34 , 108628. [ Google Scholar ] [ CrossRef ]
- Liu, G.; Park, H.S.; Pyo, H.M.; Liu, Q.; Zhou, Y. Influenza A Virus Panhandle Structure Is Directly Involved in RIG-I Activation and Interferon Induction. J. Virol. 2015 , 89 , 6067–6079. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Te Velthuis, A.J.W.; Long, J.C.; Bauer, D.L.V.; Fan, R.L.Y.; Yen, H.L.; Sharps, J.; Siegers, J.Y.; Killip, M.J.; French, H.; Oliva-Martin, M.J.; et al. Mini viral RNAs act as innate immune agonists during influenza virus infection. Nat. Microbiol. 2018 , 3 , 1234–1242. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Lee, M.K.; Kim, H.E.; Park, E.B.; Lee, J.; Kim, K.H.; Lim, K.; Yum, S.; Lee, Y.H.; Kang, S.J.; Lee, J.H.; et al. Structural features of influenza A virus panhandle RNA enabling the activation of RIG-I independently of 5′-triphosphate. Nucleic Acids Res. 2016 , 44 , 8407–8416. [ Google Scholar ] [ CrossRef ]
- Liu, G.; Lu, Y.; Thulasi Raman, S.N.; Xu, F.; Wu, Q.; Li, Z.; Brownlie, R.; Liu, Q.; Zhou, Y. Nuclear-resident RIG-I senses viral replication inducing antiviral immunity. Nat. Commun. 2018 , 9 , 3199. [ Google Scholar ] [ CrossRef ]
- Fu, Y.; Comella, N.; Tognazzi, K.; Brown, L.F.; Dvorak, H.F.; Kocher, O. Cloning of DLM-1, a novel gene that is up-regulated in activated macrophages, using RNA differential display. Gene 1999 , 240 , 157–163. [ Google Scholar ] [ CrossRef ]
- Schwartz, T.; Behlke, J.; Lowenhaupt, K.; Heinemann, U.; Rich, A. Structure of the DLM-1-Z-DNA complex reveals a conserved family of Z-DNA-binding proteins. Nat. Struct. Biol. 2001 , 8 , 761–765. [ Google Scholar ] [ CrossRef ]
- Ha, S.C.; Van Quyen, D.; Hwang, H.Y.; Oh, D.B.; Brown, B.A., 2nd; Lee, S.M.; Park, H.J.; Ahn, J.H.; Kim, K.K.; Kim, Y.G. Biochemical characterization and preliminary X-ray crystallographic study of the domains of human ZBP1 bound to left-handed Z-DNA. Biochim. Biophys. Acta 2006 , 1764 , 320–323. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Deigendesch, N.; Koch-Nolte, F.; Rothenburg, S. ZBP1 subcellular localization and association with stress granules is controlled by its Z-DNA binding domains. Nucleic Acids Res. 2006 , 34 , 5007–5020. [ Google Scholar ] [ CrossRef ]
- Takaoka, A.; Wang, Z.; Choi, M.K.; Yanai, H.; Negishi, H.; Ban, T.; Lu, Y.; Miyagishi, M.; Kodama, T.; Honda, K.; et al. DAI (DLM-1/ZBP1) is a cytosolic DNA sensor and an activator of innate immune response. Nature 2007 , 448 , 501–505. [ Google Scholar ] [ CrossRef ]
- Rebsamen, M.; Heinz, L.X.; Meylan, E.; Michallet, M.C.; Schroder, K.; Hofmann, K.; Vazquez, J.; Benedict, C.A.; Tschopp, J. DAI/ZBP1 recruits RIP1 and RIP3 through RIP homotypic interaction motifs to activate NF-kappaB. EMBO Rep. 2009 , 10 , 916–922. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Maelfait, J.; Liverpool, L.; Bridgeman, A.; Ragan, K.B.; Upton, J.W.; Rehwinkel, J. Sensing of viral and endogenous RNA by ZBP1/DAI induces necroptosis. EMBO J. 2017 , 36 , 2529–2543. [ Google Scholar ] [ CrossRef ]
- Ha, S.C.; Kim, D.; Hwang, H.Y.; Rich, A.; Kim, Y.G.; Kim, K.K. The crystal structure of the second Z-DNA binding domain of human DAI (ZBP1) in complex with Z-DNA reveals an unusual binding mode to Z-DNA. Proc. Natl. Acad. Sci. USA 2008 , 105 , 20671–20676. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Song, Q.; Fan, Y.; Zhang, H.; Wang, N. Z-DNA binding protein 1 orchestrates innate immunity and inflammatory cell death. Cytokine Growth Factor Rev. 2024 , 77 , 15–29. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Pham, H.T.; Park, M.Y.; Kim, K.K.; Kim, Y.G.; Ahn, J.H. Intracellular localization of human ZBP1: Differential regulation by the Z-DNA binding domain, Zalpha, in splice variants. Biochem. Biophys. Res. Commun. 2006 , 348 , 145–152. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Kuriakose, T.; Man, S.M.; Malireddi, R.K.; Karki, R.; Kesavardhana, S.; Place, D.E.; Neale, G.; Vogel, P.; Kanneganti, T.D. ZBP1/DAI is an innate sensor of influenza virus triggering the NLRP3 inflammasome and programmed cell death pathways. Sci. Immunol. 2016 , 1 , aag2045. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Zhang, T.; Yin, C.; Boyd, D.F.; Quarato, G.; Ingram, J.P.; Shubina, M.; Ragan, K.B.; Ishizuka, T.; Crawford, J.C.; Tummers, B.; et al. Influenza Virus Z-RNAs Induce ZBP1-Mediated Necroptosis. Cell 2020 , 180 , 1115–1129.e13. [ Google Scholar ] [ CrossRef ]
- Thapa, R.J.; Ingram, J.P.; Ragan, K.B.; Nogusa, S.; Boyd, D.F.; Benitez, A.A.; Sridharan, H.; Kosoff, R.; Shubina, M.; Landsteiner, V.J.; et al. DAI Senses Influenza A Virus Genomic RNA and Activates RIPK3-Dependent Cell Death. Cell Host Microbe 2016 , 20 , 674–681. [ Google Scholar ] [ CrossRef ]
- Kesavardhana, S.; Kuriakose, T.; Guy, C.S.; Samir, P.; Malireddi, R.K.S.; Mishra, A.; Kanneganti, T.D. ZBP1/DAI ubiquitination and sensing of influenza vRNPs activate programmed cell death. J. Exp. Med. 2017 , 214 , 2217–2229. [ Google Scholar ] [ CrossRef ]
- Kesavardhana, S.; Malireddi, R.K.S.; Burton, A.R.; Porter, S.N.; Vogel, P.; Pruett-Miller, S.M.; Kanneganti, T.D. The Zalpha2 domain of ZBP1 is a molecular switch regulating influenza-induced PANoptosis and perinatal lethality during development. J. Biol. Chem. 2020 , 295 , 8325–8330. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Steimle, V.; Otten, L.A.; Zufferey, M.; Mach, B. Complementation cloning of an MHC class II transactivator mutated in hereditary MHC class II deficiency (or bare lymphocyte syndrome). Cell 1993 , 75 , 135–146. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Harton, J.A.; Linhoff, M.W.; Zhang, J.; Ting, J.P. Cutting edge: CATERPILLER: A large family of mammalian genes containing CARD, pyrin, nucleotide-binding, and leucine-rich repeat domains. J. Immunol. 2002 , 169 , 4088–4093. [ Google Scholar ] [ CrossRef ]
- Schroder, K.; Tschopp, J. The inflammasomes. Cell 2010 , 140 , 821–832. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Blevins, H.M.; Xu, Y.; Biby, S.; Zhang, S. The NLRP3 Inflammasome Pathway: A Review of Mechanisms and Inhibitors for the Treatment of Inflammatory Diseases. Front. Aging Neurosci. 2022 , 14 , 879021. [ Google Scholar ] [ CrossRef ]
- Allen, I.C.; Scull, M.A.; Moore, C.B.; Holl, E.K.; McElvania-TeKippe, E.; Taxman, D.J.; Guthrie, E.H.; Pickles, R.J.; Ting, J.P. The NLRP3 inflammasome mediates in vivo innate immunity to influenza A virus through recognition of viral RNA. Immunity 2009 , 30 , 556–565. [ Google Scholar ] [ CrossRef ]
- Ichinohe, T.; Pang, I.K.; Iwasaki, A. Influenza virus activates inflammasomes via its intracellular M2 ion channel. Nat. Immunol. 2010 , 11 , 404–410. [ Google Scholar ] [ CrossRef ]
- Pandey, K.P.; Zhou, Y. Influenza A Virus Infection Activates NLRP3 Inflammasome through Trans-Golgi Network Dispersion. Viruses 2022 , 14 , 88. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- McAuley, J.L.; Tate, M.D.; MacKenzie-Kludas, C.J.; Pinar, A.; Zeng, W.; Stutz, A.; Latz, E.; Brown, L.E.; Mansell, A. Activation of the NLRP3 inflammasome by IAV virulence protein PB1-F2 contributes to severe pathophysiology and disease. PLoS Pathog. 2013 , 9 , e1003392. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Pinar, A.; Dowling, J.K.; Bitto, N.J.; Robertson, A.A.; Latz, E.; Stewart, C.R.; Drummond, G.R.; Cooper, M.A.; McAuley, J.L.; Tate, M.D.; et al. PB1-F2 Peptide Derived from Avian Influenza A Virus H7N9 Induces Inflammation via Activation of the NLRP3 Inflammasome. J. Biol. Chem. 2017 , 292 , 826–836. [ Google Scholar ] [ CrossRef ]
- Lee, S.; Ishitsuka, A.; Noguchi, M.; Hirohama, M.; Fujiyasu, Y.; Petric, P.P.; Schwemmle, M.; Staeheli, P.; Nagata, K.; Kawaguchi, A. Influenza restriction factor MxA functions as inflammasome sensor in the respiratory epithelium. Sci. Immunol. 2019 , 4 , eaau4643. [ Google Scholar ] [ CrossRef ]
- Motshwene, P.G.; Moncrieffe, M.C.; Grossmann, J.G.; Kao, C.; Ayaluru, M.; Sandercock, A.M.; Robinson, C.V.; Latz, E.; Gay, N.J. An oligomeric signaling platform formed by the Toll-like receptor signal transducers MyD88 and IRAK-4. J. Biol. Chem. 2009 , 284 , 25404–25411. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Vyncke, L.; Bovijn, C.; Pauwels, E.; Van Acker, T.; Ruyssinck, E.; Burg, E.; Tavernier, J.; Peelman, F. Reconstructing the TIR Side of the Myddosome: A Paradigm for TIR-TIR Interactions. Structure 2016 , 24 , 437–447. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Kawagoe, T.; Sato, S.; Matsushita, K.; Kato, H.; Matsui, K.; Kumagai, Y.; Saitoh, T.; Kawai, T.; Takeuchi, O.; Akira, S. Sequential control of Toll-like receptor-dependent responses by IRAK1 and IRAK2. Nat. Immunol. 2008 , 9 , 684–691. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Li, S.; Strelow, A.; Fontana, E.J.; Wesche, H. IRAK-4: A novel member of the IRAK family with the properties of an IRAK-kinase. Proc. Natl. Acad. Sci. USA 2002 , 99 , 5567–5572. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Wang, C.; Deng, L.; Hong, M.; Akkaraju, G.R.; Inoue, J.; Chen, Z.J. TAK1 is a ubiquitin-dependent kinase of MKK and IKK. Nature 2001 , 412 , 346–351. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Wu, C.J.; Conze, D.B.; Li, T.; Srinivasula, S.M.; Ashwell, J.D. Sensing of Lys 63-linked polyubiquitination by NEMO is a key event in NF-kappaB activation [corrected]. Nat. Cell Biol. 2006 , 8 , 398–406. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Napetschnig, J.; Wu, H. Molecular basis of NF-kappaB signaling. Annu. Rev. Biophys. 2013 , 42 , 443–468. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Kawai, T.; Sato, S.; Ishii, K.J.; Coban, C.; Hemmi, H.; Yamamoto, M.; Terai, K.; Matsuda, M.; Inoue, J.; Uematsu, S.; et al. Interferon-alpha induction through Toll-like receptors involves a direct interaction of IRF7 with MyD88 and TRAF6. Nat. Immunol. 2004 , 5 , 1061–1068. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Schoenemeyer, A.; Barnes, B.J.; Mancl, M.E.; Latz, E.; Goutagny, N.; Pitha, P.M.; Fitzgerald, K.A.; Golenbock, D.T. The interferon regulatory factor, IRF5, is a central mediator of toll-like receptor 7 signaling. J. Biol. Chem. 2005 , 280 , 17005–17012. [ Google Scholar ] [ CrossRef ]
- Takaoka, A.; Yanai, H.; Kondo, S.; Duncan, G.; Negishi, H.; Mizutani, T.; Kano, S.; Honda, K.; Ohba, Y.; Mak, T.W.; et al. Integral role of IRF-5 in the gene induction programme activated by Toll-like receptors. Nature 2005 , 434 , 243–249. [ Google Scholar ] [ CrossRef ]
- Bergstrom, B.; Aune, M.H.; Awuh, J.A.; Kojen, J.F.; Blix, K.J.; Ryan, L.; Flo, T.H.; Mollnes, T.E.; Espevik, T.; Stenvik, J. TLR8 Senses Staphylococcus aureus RNA in Human Primary Monocytes and Macrophages and Induces IFN-beta Production via a TAK1-IKKbeta-IRF5 Signaling Pathway. J. Immunol. 2015 , 195 , 1100–1111. [ Google Scholar ] [ CrossRef ]
- Alexopoulou, L.; Holt, A.C.; Medzhitov, R.; Flavell, R.A. Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature 2001 , 413 , 732–738. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Hoebe, K.; Du, X.; Georgel, P.; Janssen, E.; Tabeta, K.; Kim, S.O.; Goode, J.; Lin, P.; Mann, N.; Mudd, S.; et al. Identification of Lps2 as a key transducer of MyD88-independent TIR signalling. Nature 2003 , 424 , 743–748. [ Google Scholar ] [ CrossRef ]
- Oshiumi, H.; Matsumoto, M.; Funami, K.; Akazawa, T.; Seya, T. TICAM-1, an adaptor molecule that participates in Toll-like receptor 3-mediated interferon-beta induction. Nat. Immunol. 2003 , 4 , 161–167. [ Google Scholar ] [ CrossRef ]
- Yamamoto, M.; Sato, S.; Hemmi, H.; Hoshino, K.; Kaisho, T.; Sanjo, H.; Takeuchi, O.; Sugiyama, M.; Okabe, M.; Takeda, K.; et al. Role of adaptor TRIF in the MyD88-independent toll-like receptor signaling pathway. Science 2003 , 301 , 640–643. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Cusson-Hermance, N.; Khurana, S.; Lee, T.H.; Fitzgerald, K.A.; Kelliher, M.A. Rip1 mediates the Trif-dependent toll-like receptor 3- and 4-induced NF-kappaB activation but does not contribute to interferon regulatory factor 3 activation. J. Biol. Chem. 2005 , 280 , 36560–36566. [ Google Scholar ] [ CrossRef ]
- Meylan, E.; Burns, K.; Hofmann, K.; Blancheteau, V.; Martinon, F.; Kelliher, M.; Tschopp, J. RIP1 is an essential mediator of Toll-like receptor 3-induced NF-kappa B activation. Nat. Immunol. 2004 , 5 , 503–507. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Chen, F.; Chen, L.; Li, Y.; Sang, H.; Zhang, C.; Yuan, S.; Yang, J. TRAF3 Positively Regulates Host Innate Immune Resistance to Influenza A Virus Infection. Front. Cell. Infect. Microbiol. 2022 , 12 , 839625. [ Google Scholar ] [ CrossRef ]
- Hacker, H.; Redecke, V.; Blagoev, B.; Kratchmarova, I.; Hsu, L.C.; Wang, G.G.; Kamps, M.P.; Raz, E.; Wagner, H.; Hacker, G.; et al. Specificity in Toll-like receptor signalling through distinct effector functions of TRAF3 and TRAF6. Nature 2006 , 439 , 204–207. [ Google Scholar ] [ CrossRef ]
- Oganesyan, G.; Saha, S.K.; Guo, B.; He, J.Q.; Shahangian, A.; Zarnegar, B.; Perry, A.; Cheng, G. Critical role of TRAF3 in the Toll-like receptor-dependent and -independent antiviral response. Nature 2006 , 439 , 208–211. [ Google Scholar ] [ CrossRef ]
- Malik, G.; Zhou, Y. Innate Immune Sensing of Influenza A Virus. Viruses 2020 , 12 , 755. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Seo, S.U.; Kwon, H.J.; Song, J.H.; Byun, Y.H.; Seong, B.L.; Kawai, T.; Akira, S.; Kweon, M.N. MyD88 signaling is indispensable for primary influenza A virus infection but dispensable for secondary infection. J. Virol. 2010 , 84 , 12713–12722. [ Google Scholar ] [ CrossRef ]
- Wu, W.; Zhang, W.; Duggan, E.S.; Booth, J.L.; Zou, M.H.; Metcalf, J.P. RIG-I and TLR3 are both required for maximum interferon induction by influenza virus in human lung alveolar epithelial cells. Virology 2015 , 482 , 181–188. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Huo, C.; Jin, Y.; Zou, S.; Qi, P.; Xiao, J.; Tian, H.; Wang, M.; Hu, Y. Lethal influenza A virus preferentially activates TLR3 and triggers a severe inflammatory response. Virus Res. 2018 , 257 , 102–112. [ Google Scholar ] [ CrossRef ]
- Yong, Y.H.; Liu, S.F.; Hua, G.H.; Jia, R.M.; Gooneratne, R.; Zhao, Y.T.; Liao, M.; Ju, X.H. Goose toll-like receptor 3 (TLR3) mediated IFN-gamma and IL-6 in anti-H5N1 avian influenza virus response. Vet. Immunol. Immunopathol. 2018 , 197 , 31–38. [ Google Scholar ] [ CrossRef ]
- Park, W.J.; Han, S.H.; Kim, D.H.; Song, Y.J.; Lee, J.B.; Park, S.Y.; Song, C.S.; Lee, S.W.; Choi, I.S. Induction of IFN-beta through TLR-3- and RIG-I-Mediated Signaling Pathways in Canine Respiratory Epithelial Cells Infected with H3N2 Canine Influenza Virus. J. Microbiol. Biotechnol. 2021 , 31 , 942–948. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- de Marcken, M.; Dhaliwal, K.; Danielsen, A.C.; Gautron, A.S.; Dominguez-Villar, M. TLR7 and TLR8 activate distinct pathways in monocytes during RNA virus infection. Sci. Signal. 2019 , 12 , aaw1347. [ Google Scholar ] [ CrossRef ]
- Wies, E.; Wang, M.K.; Maharaj, N.P.; Chen, K.; Zhou, S.; Finberg, R.W.; Gack, M.U. Dephosphorylation of the RNA sensors RIG-I and MDA5 by the phosphatase PP1 is essential for innate immune signaling. Immunity 2013 , 38 , 437–449. [ Google Scholar ] [ CrossRef ]
- Nistal-Villan, E.; Gack, M.U.; Martinez-Delgado, G.; Maharaj, N.P.; Inn, K.S.; Yang, H.; Wang, R.; Aggarwal, A.K.; Jung, J.U.; Garcia-Sastre, A. Negative role of RIG-I serine 8 phosphorylation in the regulation of interferon-beta production. J. Biol. Chem. 2010 , 285 , 20252–20261. [ Google Scholar ] [ CrossRef ]
- Gack, M.U.; Nistal-Villan, E.; Inn, K.S.; Garcia-Sastre, A.; Jung, J.U. Phosphorylation-mediated negative regulation of RIG-I antiviral activity. J. Virol. 2010 , 84 , 3220–3229. [ Google Scholar ] [ CrossRef ]
- Sun, Z.; Ren, H.; Liu, Y.; Teeling, J.L.; Gu, J. Phosphorylation of RIG-I by casein kinase II inhibits its antiviral response. J. Virol. 2011 , 85 , 1036–1047. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Maharaj, N.P.; Wies, E.; Stoll, A.; Gack, M.U. Conventional protein kinase C-alpha (PKC-alpha) and PKC-beta negatively regulate RIG-I antiviral signal transduction. J. Virol. 2012 , 86 , 1358–1371. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Kowalinski, E.; Lunardi, T.; McCarthy, A.A.; Louber, J.; Brunel, J.; Grigorov, B.; Gerlier, D.; Cusack, S. Structural basis for the activation of innate immune pattern-recognition receptor RIG-I by viral RNA. Cell 2011 , 147 , 423–435. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Gee, P.; Chua, P.K.; Gevorkyan, J.; Klumpp, K.; Najera, I.; Swinney, D.C.; Deval, J. Essential role of the N-terminal domain in the regulation of RIG-I ATPase activity. J. Biol. Chem. 2008 , 283 , 9488–9496. [ Google Scholar ] [ CrossRef ]
- Jiang, F.; Ramanathan, A.; Miller, M.T.; Tang, G.Q.; Gale, M., Jr.; Patel, S.S.; Marcotrigiano, J. Structural basis of RNA recognition and activation by innate immune receptor RIG-I. Nature 2011 , 479 , 423–427. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Cui, S.; Eisenacher, K.; Kirchhofer, A.; Brzozka, K.; Lammens, A.; Lammens, K.; Fujita, T.; Conzelmann, K.K.; Krug, A.; Hopfner, K.P. The C-terminal regulatory domain is the RNA 5′-triphosphate sensor of RIG-I. Mol. Cell 2008 , 29 , 169–179. [ Google Scholar ] [ CrossRef ]
- Luo, D.; Ding, S.C.; Vela, A.; Kohlway, A.; Lindenbach, B.D.; Pyle, A.M. Structural insights into RNA recognition by RIG-I. Cell 2011 , 147 , 409–422. [ Google Scholar ] [ CrossRef ]
- Peisley, A.; Wu, B.; Yao, H.; Walz, T.; Hur, S. RIG-I forms signaling-competent filaments in an ATP-dependent, ubiquitin-independent manner. Mol. Cell 2013 , 51 , 573–583. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Luo, D.; Kohlway, A.; Vela, A.; Pyle, A.M. Visualizing the determinants of viral RNA recognition by innate immune sensor RIG-I. Structure 2012 , 20 , 1983–1988. [ Google Scholar ] [ CrossRef ]
- Gao, D.; Yang, Y.K.; Wang, R.P.; Zhou, X.; Diao, F.C.; Li, M.D.; Zhai, Z.H.; Jiang, Z.F.; Chen, D.Y. REUL is a novel E3 ubiquitin ligase and stimulator of retinoic-acid-inducible gene-I. PLoS ONE 2009 , 4 , e5760. [ Google Scholar ] [ CrossRef ]
- Oshiumi, H.; Matsumoto, M.; Hatakeyama, S.; Seya, T. Riplet/RNF135, a RING finger protein, ubiquitinates RIG-I to promote interferon-beta induction during the early phase of viral infection. J. Biol. Chem. 2009 , 284 , 807–817. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Oshiumi, H.; Miyashita, M.; Inoue, N.; Okabe, M.; Matsumoto, M.; Seya, T. The ubiquitin ligase Riplet is essential for RIG-I-dependent innate immune responses to RNA virus infection. Cell Host Microbe 2010 , 8 , 496–509. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Gack, M.U.; Shin, Y.C.; Joo, C.H.; Urano, T.; Liang, C.; Sun, L.; Takeuchi, O.; Akira, S.; Chen, Z.; Inoue, S.; et al. TRIM25 RING-finger E3 ubiquitin ligase is essential for RIG-I-mediated antiviral activity. Nature 2007 , 446 , 916–920. [ Google Scholar ] [ CrossRef ]
- Zeng, W.; Sun, L.; Jiang, X.; Chen, X.; Hou, F.; Adhikari, A.; Xu, M.; Chen, Z.J. Reconstitution of the RIG-I pathway reveals a signaling role of unanchored polyubiquitin chains in innate immunity. Cell 2010 , 141 , 315–330. [ Google Scholar ] [ CrossRef ]
- Xian, H.; Xie, W.; Yang, S.; Liu, Q.; Xia, X.; Jin, S.; Sun, T.; Cui, J. Stratified ubiquitination of RIG-I creates robust immune response and induces selective gene expression. Sci. Adv. 2017 , 3 , e1701764. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Hayman, T.J.; Hsu, A.C.; Kolesnik, T.B.; Dagley, L.F.; Willemsen, J.; Tate, M.D.; Baker, P.J.; Kershaw, N.J.; Kedzierski, L.; Webb, A.I.; et al. RIPLET, and not TRIM25, is required for endogenous RIG-I-dependent antiviral responses. Immunol. Cell Biol. 2019 , 97 , 840–852. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Jiang, X.; Kinch, L.N.; Brautigam, C.A.; Chen, X.; Du, F.; Grishin, N.V.; Chen, Z.J. Ubiquitin-induced oligomerization of the RNA sensors RIG-I and MDA5 activates antiviral innate immune response. Immunity 2012 , 36 , 959–973. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Liu, H.M.; Loo, Y.M.; Horner, S.M.; Zornetzer, G.A.; Katze, M.G.; Gale, M., Jr. The mitochondrial targeting chaperone 14-3-3epsilon regulates a RIG-I translocon that mediates membrane association and innate antiviral immunity. Cell Host Microbe 2012 , 11 , 528–537. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Horner, S.M.; Liu, H.M.; Park, H.S.; Briley, J.; Gale, M., Jr. Mitochondrial-associated endoplasmic reticulum membranes (MAM) form innate immune synapses and are targeted by hepatitis C virus. Proc. Natl. Acad. Sci. USA 2011 , 108 , 14590–14595. [ Google Scholar ] [ CrossRef ]
- Seth, R.B.; Sun, L.; Ea, C.K.; Chen, Z.J. Identification and characterization of MAVS, a mitochondrial antiviral signaling protein that activates NF-kappaB and IRF 3. Cell 2005 , 122 , 669–682. [ Google Scholar ] [ CrossRef ]
- Dixit, E.; Boulant, S.; Zhang, Y.; Lee, A.S.; Odendall, C.; Shum, B.; Hacohen, N.; Chen, Z.J.; Whelan, S.P.; Fransen, M.; et al. Peroxisomes are signaling platforms for antiviral innate immunity. Cell 2010 , 141 , 668–681. [ Google Scholar ] [ CrossRef ]
- Hou, F.; Sun, L.; Zheng, H.; Skaug, B.; Jiang, Q.X.; Chen, Z.J. MAVS forms functional prion-like aggregates to activate and propagate antiviral innate immune response. Cell 2011 , 146 , 448–461. [ Google Scholar ] [ CrossRef ]
- Michallet, M.C.; Meylan, E.; Ermolaeva, M.A.; Vazquez, J.; Rebsamen, M.; Curran, J.; Poeck, H.; Bscheider, M.; Hartmann, G.; Konig, M.; et al. TRADD protein is an essential component of the RIG-like helicase antiviral pathway. Immunity 2008 , 28 , 651–661. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Guo, B.; Cheng, G. Modulation of the interferon antiviral response by the TBK1/IKKi adaptor protein TANK. J. Biol. Chem. 2007 , 282 , 11817–11826. [ Google Scholar ] [ CrossRef ]
- Zhao, T.; Yang, L.; Sun, Q.; Arguello, M.; Ballard, D.W.; Hiscott, J.; Lin, R. The NEMO adaptor bridges the nuclear factor-kappaB and interferon regulatory factor signaling pathways. Nat. Immunol. 2007 , 8 , 592–600. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Pippig, D.A.; Hellmuth, J.C.; Cui, S.; Kirchhofer, A.; Lammens, K.; Lammens, A.; Schmidt, A.; Rothenfusser, S.; Hopfner, K.P. The regulatory domain of the RIG-I family ATPase LGP2 senses double-stranded RNA. Nucleic Acids Res. 2009 , 37 , 2014–2025. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Li, X.; Ranjith-Kumar, C.T.; Brooks, M.T.; Dharmaiah, S.; Herr, A.B.; Kao, C.; Li, P. The RIG-I-like receptor LGP2 recognizes the termini of double-stranded RNA. J. Biol. Chem. 2009 , 284 , 13881–13891. [ Google Scholar ] [ CrossRef ]
- Rothenfusser, S.; Goutagny, N.; DiPerna, G.; Gong, M.; Monks, B.G.; Schoenemeyer, A.; Yamamoto, M.; Akira, S.; Fitzgerald, K.A. The RNA helicase Lgp2 inhibits TLR-independent sensing of viral replication by retinoic acid-inducible gene-I. J. Immunol. 2005 , 175 , 5260–5268. [ Google Scholar ] [ CrossRef ]
- Rodriguez, K.R.; Bruns, A.M.; Horvath, C.M. MDA5 and LGP2: Accomplices and antagonists of antiviral signal transduction. J. Virol. 2014 , 88 , 8194–8200. [ Google Scholar ] [ CrossRef ]
- Childs, K.S.; Randall, R.E.; Goodbourn, S. LGP2 plays a critical role in sensitizing mda-5 to activation by double-stranded RNA. PLoS ONE 2013 , 8 , e64202. [ Google Scholar ] [ CrossRef ]
- Uchikawa, E.; Lethier, M.; Malet, H.; Brunel, J.; Gerlier, D.; Cusack, S. Structural Analysis of dsRNA Binding to Anti-viral Pattern Recognition Receptors LGP2 and MDA5. Mol. Cell 2016 , 62 , 586–602. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Bruns, A.M.; Leser, G.P.; Lamb, R.A.; Horvath, C.M. The innate immune sensor LGP2 activates antiviral signaling by regulating MDA5-RNA interaction and filament assembly. Mol. Cell 2014 , 55 , 771–781. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Duic, I.; Tadakuma, H.; Harada, Y.; Yamaue, R.; Deguchi, K.; Suzuki, Y.; Yoshimura, S.H.; Kato, H.; Takeyasu, K.; Fujita, T. Viral RNA recognition by LGP2 and MDA5, and activation of signaling through step-by-step conformational changes. Nucleic Acids Res. 2020 , 48 , 11664–11674. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Bruns, A.M.; Pollpeter, D.; Hadizadeh, N.; Myong, S.; Marko, J.F.; Horvath, C.M. ATP hydrolysis enhances RNA recognition and antiviral signal transduction by the innate immune sensor, laboratory of genetics and physiology 2 (LGP2). J. Biol. Chem. 2013 , 288 , 938–946. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Singh, R.; Wu, Y.; Herrero Del Valle, A.; Leigh, K.E.; Mong, S.; Cheng, M.T.K.; Ferguson, B.J.; Modis, Y. Contrasting functions of ATP hydrolysis by MDA5 and LGP2 in viral RNA sensing. J. Biol. Chem. 2024 , 300 , 105711. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Opitz, B.; Rejaibi, A.; Dauber, B.; Eckhard, J.; Vinzing, M.; Schmeck, B.; Hippenstiel, S.; Suttorp, N.; Wolff, T. IFNbeta induction by influenza A virus is mediated by RIG-I which is regulated by the viral NS1 protein. Cell Microbiol. 2007 , 9 , 930–938. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Kandasamy, M.; Suryawanshi, A.; Tundup, S.; Perez, J.T.; Schmolke, M.; Manicassamy, S.; Manicassamy, B. RIG-I Signaling Is Critical for Efficient Polyfunctional T Cell Responses during Influenza Virus Infection. PLoS Pathog. 2016 , 12 , e1005754. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Hao, W.; Wang, L.; Li, S. FKBP5 Regulates RIG-I-Mediated NF-kappaB Activation and Influenza A Virus Infection. Viruses 2020 , 12 , 672. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Sun, N.; Jiang, L.; Ye, M.; Wang, Y.; Wang, G.; Wan, X.; Zhao, Y.; Wen, X.; Liang, L.; Ma, S.; et al. TRIM35 mediates protection against influenza infection by activating TRAF3 and degrading viral PB2. Protein Cell 2020 , 11 , 894–914. [ Google Scholar ] [ CrossRef ]
- Mohamed, A.A.; Soler, S.; Wegner, J.; Bartok, E.; Stankovic, S.; Brooks, A.G.; Schlee, M. Influenza A Infection Stimulates RIG-I and Enhances Effector Function of Primary Human NK Cells. Int. J. Mol. Sci. 2023 , 24 , 12220. [ Google Scholar ] [ CrossRef ]
- Barber, M.R.; Aldridge, J.R., Jr.; Fleming-Canepa, X.; Wang, Y.D.; Webster, R.G.; Magor, K.E. Identification of avian RIG-I responsive genes during influenza infection. Mol. Immunol. 2013 , 54 , 89–97. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Blaine, A.H.; Miranzo-Navarro, D.; Campbell, L.K.; Aldridge, J.R., Jr.; Webster, R.G.; Magor, K.E. Duck TRIM27-L enhances MAVS signaling and is absent in chickens and turkeys. Mol. Immunol. 2015 , 67 , 607–615. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Shao, Q.; Xu, W.; Guo, Q.; Yan, L.; Rui, L.; Liu, J.; Zhao, Y.; Li, Z. RIG-I from waterfowl and mammals differ in their abilities to induce antiviral responses against influenza A viruses. J. Gen. Virol. 2015 , 96 , 277–287. [ Google Scholar ] [ CrossRef ]
- Wang, Z.; Ye, S.; Yao, C.; Wang, J.; Mao, J.; Xu, L.; Liu, Y.; Fu, C.; Lu, G.; Li, S. Antiviral Activity of Canine RIG-I against Canine Influenza Virus and Interactions between Canine RIG-I and CIV. Viruses 2021 , 13 , 2048. [ Google Scholar ] [ CrossRef ]
- Zhai, B.; Liu, L.; Li, X.; Lv, X.; Wu, J.; Li, J.; Lin, S.; Yin, Y.; Lan, J.; Du, J.; et al. The Variation of Duck RIG-I-Mediated Innate Immune Response Induced by Different Virulence Avian Influenza Viruses. Front. Microbiol. 2022 , 13 , 842721. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Barber, M.R.; Aldridge, J.R., Jr.; Webster, R.G.; Magor, K.E. Association of RIG-I with innate immunity of ducks to influenza. Proc. Natl. Acad. Sci. USA 2010 , 107 , 5913–5918. [ Google Scholar ] [ CrossRef ]
- Wei, Y.; Zeng, Y.; Zhang, X.; Xu, S.; Wang, Z.; Du, Y.; Zhang, B.; Lei, C.Q.; Zhu, Q. The Nucleoprotein of H7N9 Influenza Virus Positively Regulates TRAF3-Mediated Innate Signaling and Attenuates Viral Virulence in Mice. J. Virol. 2020 , 94 , JVI.01640-20. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Ranjan, P.; Singh, N.; Kumar, A.; Neerincx, A.; Kremmer, E.; Cao, W.; Davis, W.G.; Katz, J.M.; Gangappa, S.; Lin, R.; et al. NLRC5 interacts with RIG-I to induce a robust antiviral response against influenza virus infection. Eur. J. Immunol. 2015 , 45 , 758–772. [ Google Scholar ] [ CrossRef ]
- Zhao, L.; Zhu, J.; Zhou, H.; Zhao, Z.; Zou, Z.; Liu, X.; Lin, X.; Zhang, X.; Deng, X.; Wang, R.; et al. Identification of cellular microRNA-136 as a dual regulator of RIG-I-mediated innate immunity that antagonizes H5N1 IAV replication in A549 cells. Sci. Rep. 2015 , 5 , 14991. [ Google Scholar ] [ CrossRef ]
- Song, Y.; Lai, L.; Chong, Z.; He, J.; Zhang, Y.; Xue, Y.; Xie, Y.; Chen, S.; Dong, P.; Chen, L.; et al. E3 ligase FBXW7 is critical for RIG-I stabilization during antiviral responses. Nat. Commun. 2017 , 8 , 14654. [ Google Scholar ] [ CrossRef ]
- Jahan, A.S.; Biquand, E.; Munoz-Moreno, R.; Le Quang, A.; Mok, C.K.; Wong, H.H.; Teo, Q.W.; Valkenburg, S.A.; Chin, A.W.H.; Man Poon, L.L.; et al. OTUB1 Is a Key Regulator of RIG-I-Dependent Immune Signaling and Is Targeted for Proteasomal Degradation by Influenza A NS1. Cell Rep. 2020 , 30 , 1570–1584.e6. [ Google Scholar ] [ CrossRef ]
- Nunez, R.D.; Budt, M.; Saenger, S.; Paki, K.; Arnold, U.; Sadewasser, A.; Wolff, T. The RNA Helicase DDX6 Associates with RIG-I to Augment Induction of Antiviral Signaling. Int. J. Mol. Sci. 2018 , 19 , 1877. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Marcos-Villar, L.; Nistal-Villan, E.; Zamarreno, N.; Garaigorta, U.; Gastaminza, P.; Nieto, A. Interferon-β Stimulation Elicited by the Influenza Virus Is Regulated by the Histone Methylase Dot1L through the RIG-I-TRIM25 Signaling Axis. Cells 2020 , 9 , 732. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Jiang, Z.; Wei, F.; Zhang, Y.; Wang, T.; Gao, W.; Yu, S.; Sun, H.; Pu, J.; Sun, Y.; Wang, M.; et al. IFI16 directly senses viral RNA and enhances RIG-I transcription and activation to restrict influenza virus infection. Nat. Microbiol. 2021 , 6 , 932–945. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Steinberg, J.; Wadenpohl, T.; Jung, S. The Endogenous RIG-I Ligand Is Generated in Influenza A-Virus Infected Cells. Viruses 2021 , 13 , 1564. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Acharya, D.; Reis, R.; Volcic, M.; Liu, G.; Wang, M.K.; Chia, B.S.; Nchioua, R.; Gross, R.; Munch, J.; Kirchhoff, F.; et al. Actin cytoskeleton remodeling primes RIG-I-like receptor activation. Cell 2022 , 185 , 3588–3602.e21. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Hage, A.; Bharaj, P.; van Tol, S.; Giraldo, M.I.; Gonzalez-Orozco, M.; Valerdi, K.M.; Warren, A.N.; Aguilera-Aguirre, L.; Xie, X.; Widen, S.G.; et al. The RNA helicase DHX16 recognizes specific viral RNA to trigger RIG-I-dependent innate antiviral immunity. Cell Rep. 2022 , 38 , 110434. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Cheng, J.; Tao, J.; Li, B.; Shi, Y.; Liu, H. The lncRNA HCG4 regulates the RIG-I-mediated IFN production to suppress H1N1 swine influenza virus replication. Front. Microbiol. 2023 , 14 , 1324218. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Zhang, F.; Liu, S.; Qiao, Z.; Li, L.; Han, Y.; Sun, J.; Ge, C.; Zhu, J.; Li, D.; Yao, H.; et al. Housekeeping U1 snRNA facilitates antiviral innate immunity by promoting TRIM25-mediated RIG-I activation. Cell Rep. 2024 , 43 , 113945. [ Google Scholar ] [ CrossRef ]
- Choi, S.J.; Lee, H.C.; Kim, J.H.; Park, S.Y.; Kim, T.H.; Lee, W.K.; Jang, D.J.; Yoon, J.E.; Choi, Y.I.; Kim, S.; et al. HDAC6 regulates cellular viral RNA sensing by deacetylation of RIG-I. EMBO J. 2016 , 35 , 429–442. [ Google Scholar ] [ CrossRef ]
- Benitez, A.A.; Panis, M.; Xue, J.; Varble, A.; Shim, J.V.; Frick, A.L.; Lopez, C.B.; Sachs, D.; tenOever, B.R. In Vivo RNAi Screening Identifies MDA5 as a Significant Contributor to the Cellular Defense against Influenza A Virus. Cell Rep. 2015 , 11 , 1714–1726. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Fu, C.; Ye, S.; Liu, Y.; Li, S. Role of CARD Region of MDA5 Gene in Canine Influenza Virus Infection. Viruses 2020 , 12 , 307. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Shao, Q.; Fu, F.; Zhu, P.; Yu, X.; Wang, J.; Wang, Z.; Ma, J.; Wang, H.; Yan, Y.; Cheng, Y.; et al. Pigeon MDA5 inhibits viral replication by triggering antiviral innate immunity. Poult. Sci. 2023 , 102 , 102954. [ Google Scholar ] [ CrossRef ]
- Liniger, M.; Summerfield, A.; Zimmer, G.; McCullough, K.C.; Ruggli, N. Chicken cells sense influenza A virus infection through MDA5 and CARDIF signaling involving LGP2. J. Virol. 2012 , 86 , 705–717. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Hayashi, T.; Watanabe, C.; Suzuki, Y.; Tanikawa, T.; Uchida, Y.; Saito, T. Chicken MDA5 senses short double-stranded RNA with implications for antiviral response against avian influenza viruses in chicken. J. Innate Immun. 2014 , 6 , 58–71. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Malur, M.; Gale, M., Jr.; Krug, R.M. LGP2 downregulates interferon production during infection with seasonal human influenza A viruses that activate interferon regulatory factor 3. J. Virol. 2012 , 86 , 10733–10738. [ Google Scholar ] [ CrossRef ]
- Si-Tahar, M.; Blanc, F.; Furio, L.; Chopy, D.; Balloy, V.; Lafon, M.; Chignard, M.; Fiette, L.; Langa, F.; Charneau, P.; et al. Protective role of LGP2 in influenza virus pathogenesis. J. Infect. Dis. 2014 , 210 , 214–223. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Wang, Z.; Choi, M.K.; Ban, T.; Yanai, H.; Negishi, H.; Lu, Y.; Tamura, T.; Takaoka, A.; Nishikura, K.; Taniguchi, T. Regulation of innate immune responses by DAI (DLM-1/ZBP1) and other DNA-sensing molecules. Proc. Natl. Acad. Sci. USA 2008 , 105 , 5477–5482. [ Google Scholar ] [ CrossRef ]
- Upton, J.W.; Kaiser, W.J.; Mocarski, E.S. DAI/ZBP1/DLM-1 complexes with RIP3 to mediate virus-induced programmed necrosis that is targeted by murine cytomegalovirus vIRA. Cell Host Microbe 2012 , 11 , 290–297. [ Google Scholar ] [ CrossRef ]
- Wang, Y.; Hao, Q.; Florence, J.M.; Jung, B.G.; Kurdowska, A.K.; Samten, B.; Idell, S.; Tang, H. Influenza Virus Infection Induces ZBP1 Expression and Necroptosis in Mouse Lungs. Front. Cell. Infect. Microbiol. 2019 , 9 , 286. [ Google Scholar ] [ CrossRef ]
- Wang, X.; Xiong, J.; Zhou, D.; Zhang, S.; Wang, L.; Tian, Q.; Li, C.; Liu, J.; Wu, Y.; Li, J.; et al. TRIM34 modulates influenza virus-activated programmed cell death by targeting Z-DNA-binding protein 1 for K63-linked polyubiquitination. J. Biol. Chem. 2022 , 298 , 101611. [ Google Scholar ] [ CrossRef ]
- Nogusa, S.; Thapa, R.J.; Dillon, C.P.; Liedmann, S.; Oguin, T.H., 3rd; Ingram, J.P.; Rodriguez, D.A.; Kosoff, R.; Sharma, S.; Sturm, O.; et al. RIPK3 Activates Parallel Pathways of MLKL-Driven Necroptosis and FADD-Mediated Apoptosis to Protect against Influenza A Virus. Cell Host Microbe 2016 , 20 , 13–24. [ Google Scholar ] [ CrossRef ]
- Zheng, M.; Karki, R.; Vogel, P.; Kanneganti, T.D. Caspase-6 Is a Key Regulator of Innate Immunity, Inflammasome Activation, and Host Defense. Cell 2020 , 181 , 674–687.e13. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Malireddi, R.K.S.; Sharma, B.R.; Bynigeri, R.R.; Wang, Y.; Lu, J.; Kanneganti, T.D. ZBP1 Drives IAV-Induced NLRP3 Inflammasome Activation and Lytic Cell Death, PANoptosis, Independent of the Necroptosis Executioner MLKL. Viruses 2023 , 15 , 2141. [ Google Scholar ] [ CrossRef ]
- Lei, X.; Chen, Y.; Lien, E.; Fitzgerald, K.A. MLKL-Driven Inflammasome Activation and Caspase-8 Mediate Inflammatory Cell Death in Influenza A Virus Infection. mBio 2023 , 14 , e0011023. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int. J. Mol. Sci. 2019 , 20 , 3328. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Wan, P.; Zhang, S.; Ruan, Z.; Liu, X.; Yang, G.; Jia, Y.; Li, Y.; Pan, P.; Wang, W.; Li, G.; et al. AP-1 signaling pathway promotes pro-IL-1beta transcription to facilitate NLRP3 inflammasome activation upon influenza A virus infection. Virulence 2022 , 13 , 502–513. [ Google Scholar ] [ CrossRef ]
- Ichinohe, T.; Lee, H.K.; Ogura, Y.; Flavell, R.; Iwasaki, A. Inflammasome recognition of influenza virus is essential for adaptive immune responses. J. Exp. Med. 2009 , 206 , 79–87. [ Google Scholar ] [ CrossRef ]
- Christgen, S.; Place, D.E.; Zheng, M.; Briard, B.; Yamamoto, M.; Kanneganti, T.D. The IFN-inducible GTPase IRGB10 regulates viral replication and inflammasome activation during influenza A virus infection in mice. Eur. J. Immunol. 2022 , 52 , 285–296. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Pothlichet, J.; Meunier, I.; Davis, B.K.; Ting, J.P.; Skamene, E.; von Messling, V.; Vidal, S.M. Type I IFN triggers RIG-I/TLR3/NLRP3-dependent inflammasome activation in influenza A virus infected cells. PLoS Pathog. 2013 , 9 , e1003256. [ Google Scholar ] [ CrossRef ]
- Wang, X.; Jiang, W.; Yan, Y.; Gong, T.; Han, J.; Tian, Z.; Zhou, R. RNA viruses promote activation of the NLRP3 inflammasome through a RIP1-RIP3-DRP1 signaling pathway. Nat. Immunol. 2014 , 15 , 1126–1133. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Park, H.S.; Liu, G.; Liu, Q.; Zhou, Y. Swine Influenza Virus Induces RIPK1/DRP1-Mediated Interleukin-1 Beta Production. Viruses 2018 , 10 , 419. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Kesavardhana, S.; Samir, P.; Zheng, M.; Malireddi, R.K.S.; Karki, R.; Sharma, B.R.; Place, D.E.; Briard, B.; Vogel, P.; Kanneganti, T.D. DDX3X coordinates host defense against influenza virus by activating the NLRP3 inflammasome and type I interferon response. J. Biol. Chem. 2021 , 296 , 100579. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Park, H.S.; Lu, Y.; Pandey, K.; Liu, G.; Zhou, Y. NLRP3 Inflammasome Activation Enhanced by TRIM25 is Targeted by the NS1 Protein of 2009 Pandemic Influenza A Virus. Front. Microbiol. 2021 , 12 , 778950. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Ichinohe, T.; Yamazaki, T.; Koshiba, T.; Yanagi, Y. Mitochondrial protein mitofusin 2 is required for NLRP3 inflammasome activation after RNA virus infection. Proc. Natl. Acad. Sci. USA 2013 , 110 , 17963–17968. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Tamura, T.; Yanai, H.; Savitsky, D.; Taniguchi, T. The IRF family transcription factors in immunity and oncogenesis. Annu. Rev. Immunol. 2008 , 26 , 535–584. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Takahasi, K.; Suzuki, N.N.; Horiuchi, M.; Mori, M.; Suhara, W.; Okabe, Y.; Fukuhara, Y.; Terasawa, H.; Akira, S.; Fujita, T.; et al. X-ray crystal structure of IRF-3 and its functional implications. Nat. Struct. Biol. 2003 , 10 , 922–927. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Wang, Z.; Ji, J.; Peng, D.; Ma, F.; Cheng, G.; Qin, F.X. Complex Regulation Pattern of IRF3 Activation Revealed by a Novel Dimerization Reporter System. J. Immunol. 2016 , 196 , 4322–4330. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Jing, T.; Zhao, B.; Xu, P.; Gao, X.; Chi, L.; Han, H.; Sankaran, B.; Li, P. The Structural Basis of IRF-3 Activation upon Phosphorylation. J. Immunol. 2020 , 205 , 1886–1896. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Al Hamrashdi, M.; Brady, G. Regulation of IRF3 activation in human antiviral signaling pathways. Biochem. Pharmacol. 2022 , 200 , 115026. [ Google Scholar ] [ CrossRef ]
- Panne, D. The enhanceosome. Curr. Opin. Struct. Biol. 2008 , 18 , 236–242. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Hatesuer, B.; Hoang, H.T.; Riese, P.; Trittel, S.; Gerhauser, I.; Elbahesh, H.; Geffers, R.; Wilk, E.; Schughart, K. Deletion of Irf3 and Irf7 Genes in Mice Results in Altered Interferon Pathway Activation and Granulocyte-Dominated Inflammatory Responses to Influenza A Infection. J. Innate Immun. 2017 , 9 , 145–161. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Chen, X.; Zhou, L.; Peng, N.; Yu, H.; Li, M.; Cao, Z.; Lin, Y.; Wang, X.; Li, Q.; Wang, J.; et al. MicroRNA-302a suppresses influenza A virus-stimulated interferon regulatory factor-5 expression and cytokine storm induction. J. Biol. Chem. 2017 , 292 , 21291–21303. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Irving, A.T.; Zhang, Q.; Kong, P.S.; Luko, K.; Rozario, P.; Wen, M.; Zhu, F.; Zhou, P.; Ng, J.H.J.; Sobota, R.M.; et al. Interferon Regulatory Factors IRF1 and IRF7 Directly Regulate Gene Expression in Bats in Response to Viral Infection. Cell Rep. 2020 , 33 , 108345. [ Google Scholar ] [ CrossRef ]
- Wu, W.; Zhang, W.; Tian, L.; Brown, B.R.; Walters, M.S.; Metcalf, J.P. IRF7 Is Required for the Second Phase Interferon Induction during Influenza Virus Infection in Human Lung Epithelia. Viruses 2020 , 12 , 377. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Tuerxun, W.; Wang, Y.; Cui, C.; Yang, L.; Wang, S.; Yu, Y.; Wang, L. Expression pattern of the interferon regulatory factor family members in influenza virus induced local and systemic inflammatory responses. Clin. Immunol. 2020 , 217 , 108469. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Kim, T.H.; Zhou, H. Functional Analysis of Chicken IRF7 in Response to dsRNA Analog Poly(I:C) by Integrating Overexpression and Knockdown. PLoS ONE 2015 , 10 , e0133450. [ Google Scholar ] [ CrossRef ]
- Crotta, S.; Davidson, S.; Mahlakoiv, T.; Desmet, C.J.; Buckwalter, M.R.; Albert, M.L.; Staeheli, P.; Wack, A. Type I and type III interferons drive redundant amplification loops to induce a transcriptional signature in influenza-infected airway epithelia. PLoS Pathog. 2013 , 9 , e1003773. [ Google Scholar ] [ CrossRef ]
- Yap, G.L.R.; Sachaphibulkij, K.; Foo, S.L.; Cui, J.; Fairhurst, A.M.; Lim, L.H.K. Annexin-A1 promotes RIG-I-dependent signaling and apoptosis via regulation of the IRF3-IFNAR-STAT1-IFIT1 pathway in A549 lung epithelial cells. Cell Death Dis. 2020 , 11 , 463. [ Google Scholar ] [ CrossRef ]
- Chen, Y.; Lei, X.; Jiang, Z.; Fitzgerald, K.A. Cellular nucleic acid-binding protein is essential for type I interferon-mediated immunity to RNA virus infection. Proc. Natl. Acad. Sci. USA 2021 , 118 , e2100383118. [ Google Scholar ] [ CrossRef ]
- Qian, W.; Wei, X.; Li, Y.; Guo, K.; Zou, Z.; Zhou, H.; Jin, M. Duck interferon regulatory factor 1 acts as a positive regulator in duck innate antiviral response. Dev. Comp. Immunol. 2018 , 78 , 1–13. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Kuriakose, T.; Zheng, M.; Neale, G.; Kanneganti, T.D. IRF1 Is a Transcriptional Regulator of ZBP1 Promoting NLRP3 Inflammasome Activation and Cell Death during Influenza Virus Infection. J. Immunol. 2018 , 200 , 1489–1495. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Natoli, G.; Saccani, S.; Bosisio, D.; Marazzi, I. Interactions of NF-kappaB with chromatin: The art of being at the right place at the right time. Nat. Immunol. 2005 , 6 , 439–445. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Hayden, M.S.; Ghosh, S. Shared principles in NF-kappaB signaling. Cell 2008 , 132 , 344–362. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Taniguchi, K.; Karin, M. NF-kappaB, inflammation, immunity and cancer: Coming of age. Nat. Rev. Immunol. 2018 , 18 , 309–324. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Zhang, L.; Ye, X.; Liu, Y.; Zhang, Z.; Xia, X.; Dong, S. Research progress on the effect of traditional Chinese medicine on the activation of PRRs-mediated NF-kappaB signaling pathway to inhibit influenza pneumonia. Front. Pharmacol. 2023 , 14 , 1132388. [ Google Scholar ] [ CrossRef ]
- Reis, A.L.; McCauley, J.W. The influenza virus protein PB1-F2 interacts with IKKbeta and modulates NF-kappaB signalling. PLoS ONE 2013 , 8 , e63852. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Munir, M.; Embry, A.; Doench, J.G.; Heaton, N.S.; Wilen, C.B.; Orchard, R.C. Genome-wide CRISPR activation screen identifies JADE3 as an antiviral activator of NF-kB-dependent IFITM3 expression. J. Biol. Chem. 2024 , 300 , 107153. [ Google Scholar ] [ CrossRef ]
- Rashid, F.; Xie, Z.; Li, M.; Xie, Z.; Luo, S.; Xie, L. Roles and functions of IAV proteins in host immune evasion. Front. Immunol. 2023 , 14 , 1323560. [ Google Scholar ] [ CrossRef ]
- Killip, M.J.; Fodor, E.; Randall, R.E. Influenza virus activation of the interferon system. Virus Res. 2015 , 209 , 11–22. [ Google Scholar ] [ CrossRef ]
- Husain, M. Influenza Virus Host Restriction Factors: The ISGs and Non-ISGs. Pathogens 2024 , 13 , 127. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Koerner, I.; Kochs, G.; Kalinke, U.; Weiss, S.; Staeheli, P. Protective role of beta interferon in host defense against influenza A virus. J. Virol. 2007 , 81 , 2025–2030. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Mordstein, M.; Kochs, G.; Dumoutier, L.; Renauld, J.C.; Paludan, S.R.; Klucher, K.; Staeheli, P. Interferon-lambda contributes to innate immunity of mice against influenza A virus but not against hepatotropic viruses. PLoS Pathog. 2008 , 4 , e1000151. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Jewell, N.A.; Cline, T.; Mertz, S.E.; Smirnov, S.V.; Flano, E.; Schindler, C.; Grieves, J.L.; Durbin, R.K.; Kotenko, S.V.; Durbin, J.E. Lambda interferon is the predominant interferon induced by influenza A virus infection in vivo. J. Virol. 2010 , 84 , 11515–11522. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Galani, I.E.; Triantafyllia, V.; Eleminiadou, E.E.; Koltsida, O.; Stavropoulos, A.; Manioudaki, M.; Thanos, D.; Doyle, S.E.; Kotenko, S.V.; Thanopoulou, K.; et al. Interferon-lambda Mediates Non-redundant Front-Line Antiviral Protection against Influenza Virus Infection without Compromising Host Fitness. Immunity 2017 , 46 , 875–890.e6. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Philips, R.L.; Wang, Y.; Cheon, H.; Kanno, Y.; Gadina, M.; Sartorelli, V.; Horvath, C.M.; Darnell, J.E., Jr.; Stark, G.R.; O’Shea, J.J. The JAK-STAT pathway at 30: Much learned, much more to do. Cell 2022 , 185 , 3857–3876. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Morris, R.; Kershaw, N.J.; Babon, J.J. The molecular details of cytokine signaling via the JAK/STAT pathway. Protein Sci. 2018 , 27 , 1984–2009. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Stark, G.R.; Darnell, J.E., Jr. The JAK-STAT pathway at twenty. Immunity 2012 , 36 , 503–514. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Rengachari, S.; Groiss, S.; Devos, J.M.; Caron, E.; Grandvaux, N.; Panne, D. Structural basis of STAT2 recognition by IRF9 reveals molecular insights into ISGF3 function. Proc. Natl. Acad. Sci. USA 2018 , 115 , E601–E609. [ Google Scholar ] [ CrossRef ]
- Mahony, R.; Gargan, S.; Roberts, K.L.; Bourke, N.; Keating, S.E.; Bowie, A.G.; O’Farrelly, C.; Stevenson, N.J. A novel anti-viral role for STAT3 in IFN-alpha signalling responses. Cell. Mol. Life Sci. 2017 , 74 , 1755–1764. [ Google Scholar ] [ CrossRef ]
- Liu, S.; Liao, Y.; Chen, B.; Chen, Y.; Yu, Z.; Wei, H.; Zhang, L.; Huang, S.; Rothman, P.B.; Gao, G.F.; et al. Critical role of Syk-dependent STAT1 activation in innate antiviral immunity. Cell Rep. 2021 , 34 , 108627. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Li, X.; Liu, S.; Rai, K.R.; Zhou, W.; Wang, S.; Chi, X.; Guo, G.; Chen, J.L.; Liu, S. Initial activation of STAT2 induced by IAV infection is critical for innate antiviral immunity. Front. Immunol. 2022 , 13 , 960544. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Liu, S.; Liu, S.; Yu, Z.; Zhou, W.; Zheng, M.; Gu, R.; Hong, J.; Yang, Z.; Chi, X.; Guo, G.; et al. STAT3 regulates antiviral immunity by suppressing excessive interferon signaling. Cell Rep. 2023 , 42 , 112806. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Hermesh, T.; Moran, T.M.; Jain, D.; Lopez, C.B. Granulocyte colony-stimulating factor protects mice during respiratory virus infections. PLoS ONE 2012 , 7 , e37334. [ Google Scholar ] [ CrossRef ]
- Lauder, S.N.; Jones, E.; Smart, K.; Bloom, A.; Williams, A.S.; Hindley, J.P.; Ondondo, B.; Taylor, P.R.; Clement, M.; Fielding, C.; et al. Interleukin-6 limits influenza-induced inflammation and protects against fatal lung pathology. Eur. J. Immunol. 2013 , 43 , 2613–2625. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Liu, B.; Mori, I.; Hossain, M.J.; Dong, L.; Takeda, K.; Kimura, Y. Interleukin-18 improves the early defence system against influenza virus infection by augmenting natural killer cell-mediated cytotoxicity. J. Gen. Virol. 2004 , 85 , 423–428. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Niu, J.; Wu, S.; Chen, M.; Xu, K.; Guo, Q.; Lu, A.; Zhao, L.; Sun, B.; Meng, G. Hyperactivation of the NLRP3 inflammasome protects mice against influenza A virus infection via IL-1beta mediated neutrophil recruitment. Cytokine 2019 , 120 , 115–124. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Denton, A.E.; Doherty, P.C.; Turner, S.J.; La Gruta, N.L. IL-18, but not IL-12, is required for optimal cytokine production by influenza virus-specific CD8+ T cells. Eur. J. Immunol. 2007 , 37 , 368–375. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Schmitz, N.; Kurrer, M.; Bachmann, M.F.; Kopf, M. Interleukin-1 is responsible for acute lung immunopathology but increases survival of respiratory influenza virus infection. J. Virol. 2005 , 79 , 6441–6448. [ Google Scholar ] [ CrossRef ]
- Pang, I.K.; Ichinohe, T.; Iwasaki, A. IL-1R signaling in dendritic cells replaces pattern-recognition receptors in promoting CD8(+) T cell responses to influenza A virus. Nat. Immunol. 2013 , 14 , 246–253. [ Google Scholar ] [ CrossRef ]
- Ji, S.; Dai, M.Y.; Huang, Y.; Ren, X.C.; Jiang, M.L.; Qiao, J.P.; Zhang, W.Y.; Xu, Y.H.; Shen, J.L.; Zhang, R.Q.; et al. Influenza a virus triggers acute exacerbation of chronic obstructive pulmonary disease by increasing proinflammatory cytokines secretion via NLRP3 inflammasome activation. J. Inflamm. 2022 , 19 , 8. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Ren, R.; Wu, S.; Cai, J.; Yang, Y.; Ren, X.; Feng, Y.; Chen, L.; Qin, B.; Xu, C.; Yang, H.; et al. The H7N9 influenza A virus infection results in lethal inflammation in the mammalian host via the NLRP3-caspase-1 inflammasome. Sci. Rep. 2017 , 7 , 7625. [ Google Scholar ] [ CrossRef ]
- Rosli, S.; Harpur, C.M.; Lam, M.; West, A.C.; Hodges, C.; Mansell, A.; Lawlor, K.E.; Tate, M.D. Gasdermin D promotes hyperinflammation and immunopathology during severe influenza A virus infection. Cell Death Dis. 2023 , 14 , 727. [ Google Scholar ] [ CrossRef ]
- Ji, Z.X.; Wang, X.Q.; Liu, X.F. NS1: A Key Protein in the “Game” Between Influenza A Virus and Host in Innate Immunity. Front. Cell. Infect. Microbiol. 2021 , 11 , 670177. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Zhang, L.; Wang, J.; Munoz-Moreno, R.; Kim, M.; Sakthivel, R.; Mo, W.; Shao, D.; Anantharaman, A.; Garcia-Sastre, A.; Conrad, N.K.; et al. Influenza Virus NS1 Protein-RNA Interactome Reveals Intron Targeting. J. Virol. 2018 , 92 , JVI.01634-18. [ Google Scholar ] [ CrossRef ]
- Kumari, R.; Guo, Z.; Kumar, A.; Wiens, M.; Gangappa, S.; Katz, J.M.; Cox, N.J.; Lal, R.B.; Sarkar, D.; Fisher, P.B.; et al. Influenza virus NS1-C/EBPbeta gene regulatory complex inhibits RIG-I transcription. Antivir. Res. 2020 , 176 , 104747. [ Google Scholar ] [ CrossRef ]
- Sun, X.; Feng, W.; Guo, Y.; Wang, Q.; Dong, C.; Zhang, M.; Guan, Z.; Duan, M. MCPIP1 attenuates the innate immune response to influenza A virus by suppressing RIG-I expression in lung epithelial cells. J. Med. Virol. 2018 , 90 , 204–211. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Guo, Z.; Chen, L.M.; Zeng, H.; Gomez, J.A.; Plowden, J.; Fujita, T.; Katz, J.M.; Donis, R.O.; Sambhara, S. NS1 protein of influenza A virus inhibits the function of intracytoplasmic pathogen sensor, RIG-I. Am. J. Respir. Cell Mol. Biol. 2007 , 36 , 263–269. [ Google Scholar ] [ CrossRef ]
- Mibayashi, M.; Martinez-Sobrido, L.; Loo, Y.M.; Cardenas, W.B.; Gale, M., Jr.; Garcia-Sastre, A. Inhibition of retinoic acid-inducible gene I-mediated induction of beta interferon by the NS1 protein of influenza A virus. J. Virol. 2007 , 81 , 514–524. [ Google Scholar ] [ CrossRef ]
- Jureka, A.S.; Kleinpeter, A.B.; Cornilescu, G.; Cornilescu, C.C.; Petit, C.M. Structural Basis for a Novel Interaction between the NS1 Protein Derived from the 1918 Influenza Virus and RIG-I. Structure 2015 , 23 , 2001–2010. [ Google Scholar ] [ CrossRef ]
- Jureka, A.S.; Kleinpeter, A.B.; Tipper, J.L.; Harrod, K.S.; Petit, C.M. The influenza NS1 protein modulates RIG-I activation via a strain-specific direct interaction with the second CARD of RIG-I. J. Biol. Chem. 2020 , 295 , 1153–1164. [ Google Scholar ] [ CrossRef ]
- Rajsbaum, R.; Albrecht, R.A.; Wang, M.K.; Maharaj, N.P.; Versteeg, G.A.; Nistal-Villan, E.; Garcia-Sastre, A.; Gack, M.U. Species-specific inhibition of RIG-I ubiquitination and IFN induction by the influenza A virus NS1 protein. PLoS Pathog. 2012 , 8 , e1003059. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Gack, M.U.; Albrecht, R.A.; Urano, T.; Inn, K.S.; Huang, I.C.; Carnero, E.; Farzan, M.; Inoue, S.; Jung, J.U.; Garcia-Sastre, A. Influenza A virus NS1 targets the ubiquitin ligase TRIM25 to evade recognition by the host viral RNA sensor RIG-I. Cell Host Microbe 2009 , 5 , 439–449. [ Google Scholar ] [ CrossRef ]
- Koliopoulos, M.G.; Lethier, M.; van der Veen, A.G.; Haubrich, K.; Hennig, J.; Kowalinski, E.; Stevens, R.V.; Martin, S.R.; Reis e Sousa, C.; Cusack, S.; et al. Molecular mechanism of influenza A NS1-mediated TRIM25 recognition and inhibition. Nat. Commun. 2018 , 9 , 1820. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Evseev, D.; Miranzo-Navarro, D.; Fleming-Canepa, X.; Webster, R.G.; Magor, K.E. Avian Influenza NS1 Proteins Inhibit Human, but Not Duck, RIG-I Ubiquitination and Interferon Signaling. J. Virol. 2022 , 96 , e0077622. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Wang, T.; Wei, F.; Jiang, Z.; Song, J.; Li, C.; Liu, J. Influenza virus NS1 interacts with 14-3-3epsilon to antagonize the production of RIG-I-mediated type I interferons. Virology 2022 , 574 , 47–56. [ Google Scholar ] [ CrossRef ]
- Qian, W.; Wei, X.; Guo, K.; Li, Y.; Lin, X.; Zou, Z.; Zhou, H.; Jin, M. The C-Terminal Effector Domain of Non-Structural Protein 1 of Influenza A Virus Blocks IFN-beta Production by Targeting TNF Receptor-Associated Factor 3. Front. Immunol. 2017 , 8 , 779. [ Google Scholar ] [ CrossRef ]
- Lin, C.Y.; Shih, M.C.; Chang, H.C.; Lin, K.J.; Chen, L.F.; Huang, S.W.; Yang, M.L.; Ma, S.K.; Shiau, A.L.; Wang, J.R.; et al. Influenza a virus NS1 resembles a TRAF3-interacting motif to target the RNA sensing-TRAF3-type I IFN axis and impair antiviral innate immunity. J. Biomed. Sci. 2021 , 28 , 66. [ Google Scholar ] [ CrossRef ]
- Chen, W.; Calvo, P.A.; Malide, D.; Gibbs, J.; Schubert, U.; Bacik, I.; Basta, S.; O’Neill, R.; Schickli, J.; Palese, P.; et al. A novel influenza A virus mitochondrial protein that induces cell death. Nat. Med. 2001 , 7 , 1306–1312. [ Google Scholar ] [ CrossRef ]
- Yoshizumi, T.; Ichinohe, T.; Sasaki, O.; Otera, H.; Kawabata, S.; Mihara, K.; Koshiba, T. Influenza A virus protein PB1-F2 translocates into mitochondria via Tom40 channels and impairs innate immunity. Nat. Commun. 2014 , 5 , 4713. [ Google Scholar ] [ CrossRef ]
- Dudek, S.E.; Wixler, L.; Nordhoff, C.; Nordmann, A.; Anhlan, D.; Wixler, V.; Ludwig, S. The influenza virus PB1-F2 protein has interferon antagonistic activity. Biol. Chem. 2011 , 392 , 1135–1144. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Varga, Z.T.; Ramos, I.; Hai, R.; Schmolke, M.; Garcia-Sastre, A.; Fernandez-Sesma, A.; Palese, P. The influenza virus protein PB1-F2 inhibits the induction of type I interferon at the level of the MAVS adaptor protein. PLoS Pathog. 2011 , 7 , e1002067. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Xiao, Y.; Evseev, D.; Stevens, C.A.; Moghrabi, A.; Miranzo-Navarro, D.; Fleming-Canepa, X.; Tetrault, D.G.; Magor, K.E. Influenza PB1-F2 Inhibits Avian MAVS Signaling. Viruses 2020 , 12 , 409. [ Google Scholar ] [ CrossRef ]
- Varga, Z.T.; Grant, A.; Manicassamy, B.; Palese, P. Influenza virus protein PB1-F2 inhibits the induction of type I interferon by binding to MAVS and decreasing mitochondrial membrane potential. J. Virol. 2012 , 86 , 8359–8366. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Wang, R.; Zhu, Y.; Ren, C.; Yang, S.; Tian, S.; Chen, H.; Jin, M.; Zhou, H. Influenza A virus protein PB1-F2 impairs innate immunity by inducing mitophagy. Autophagy 2021 , 17 , 496–511. [ Google Scholar ] [ CrossRef ]
- Graef, K.M.; Vreede, F.T.; Lau, Y.F.; McCall, A.W.; Carr, S.M.; Subbarao, K.; Fodor, E. The PB2 subunit of the influenza virus RNA polymerase affects virulence by interacting with the mitochondrial antiviral signaling protein and inhibiting expression of beta interferon. J. Virol. 2010 , 84 , 8433–8445. [ Google Scholar ] [ CrossRef ]
- Patel, D.; Schultz, L.W.; Umland, T.C. Influenza A polymerase subunit PB2 possesses overlapping binding sites for polymerase subunit PB1 and human MAVS proteins. Virus Res. 2013 , 172 , 75–80. [ Google Scholar ] [ CrossRef ]
- Zeng, Y.; Xu, S.; Wei, Y.; Zhang, X.; Wang, Q.; Jia, Y.; Wang, W.; Han, L.; Chen, Z.; Wang, Z.; et al. The PB1 protein of influenza A virus inhibits the innate immune response by targeting MAVS for NBR1-mediated selective autophagic degradation. PLoS Pathog. 2021 , 17 , e1009300. [ Google Scholar ] [ CrossRef ]
- Liedmann, S.; Hrincius, E.R.; Guy, C.; Anhlan, D.; Dierkes, R.; Carter, R.; Wu, G.; Staeheli, P.; Green, D.R.; Wolff, T.; et al. Viral suppressors of the RIG-I-mediated interferon response are pre-packaged in influenza virions. Nat. Commun. 2014 , 5 , 5645. [ Google Scholar ] [ CrossRef ]
- Soonthornvacharin, S.; Rodriguez-Frandsen, A.; Zhou, Y.; Galvez, F.; Huffmaster, N.J.; Tripathi, S.; Balasubramaniam, V.R.; Inoue, A.; de Castro, E.; Moulton, H.; et al. Systems-based analysis of RIG-I-dependent signalling identifies KHSRP as an inhibitor of RIG-I receptor activation. Nat. Microbiol. 2017 , 2 , 17022. [ Google Scholar ] [ CrossRef ]
- Vogel, O.A.; Han, J.; Liang, C.Y.; Manicassamy, S.; Perez, J.T.; Manicassamy, B. The p150 Isoform of ADAR1 Blocks Sustained RLR signaling and Apoptosis during Influenza Virus Infection. PLoS Pathog. 2020 , 16 , e1008842. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Jiang, J.; Li, Y.; Sun, Z.; Gong, L.; Li, X.; Shi, F.; Yao, J.; Meng, Y.; Meng, X.; Zhang, Q.; et al. LncNSPL facilitates influenza A viral immune escape by restricting TRIM25-mediated K63-linked RIG-I ubiquitination. iScience 2022 , 25 , 104607. [ Google Scholar ] [ CrossRef ]
- Zhao, L.; Zhang, X.; Wu, Z.; Huang, K.; Sun, X.; Chen, H.; Jin, M. The Downregulation of MicroRNA hsa-miR-340-5p in IAV-Infected A549 Cells Suppresses Viral Replication by Targeting RIG-I and OAS2. Mol. Ther. Nucleic Acids 2019 , 14 , 509–519. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Shi, Q.; Li, G.; Dou, S.; Tang, L.; Hou, C.; Wang, Z.; Gao, Y.; Gao, Z.; Hao, Y.; Mo, R.; et al. Negative Regulation of RIG-I by Tim-3 Promotes H1N1 Infection. Immunol. Investig. 2023 , 52 , 1–19. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Hussain, M.; Ahmed, F.; Henzeler, B.; Husain, M. Anti-microbial host factor HDAC6 is antagonised by the influenza A virus through host caspases and viral PA. FEBS J. 2023 , 290 , 2744–2759. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Huang, K.; Zhang, Y.; Gong, W.; Yang, Y.; Jiang, L.; Zhao, L.; Yang, Y.; Wei, Y.; Li, C.; He, X.; et al. PGRMC1 Exerts Its Function of Anti-Influenza Virus in the Central Nervous System. Microbiol. Spectr. 2021 , 9 , e0073421. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Talon, J.; Horvath, C.M.; Polley, R.; Basler, C.F.; Muster, T.; Palese, P.; Garcia-Sastre, A. Activation of interferon regulatory factor 3 is inhibited by the influenza A virus NS1 protein. J. Virol. 2000 , 74 , 7989–7996. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Zu, S.; Xue, Q.; He, Z.; Shi, C.; Wu, W.; Zhang, J.; Li, W.; Huang, J.; Jiao, P.; Liao, M. Duck PIAS2 negatively regulates RIG-I mediated IFN-beta production by interacting with IRF7. Dev. Comp. Immunol. 2020 , 108 , 103664. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Zhang, B.; Liu, M.; Huang, J.; Zeng, Q.; Zhu, Q.; Xu, S.; Chen, H. H1N1 Influenza A Virus Protein NS2 Inhibits Innate Immune Response by Targeting IRF7. Viruses 2022 , 14 , 2411. [ Google Scholar ] [ CrossRef ]
- Yi, C.; Zhao, Z.; Wang, S.; Sun, X.; Zhang, D.; Sun, X.; Zhang, A.; Jin, M. Influenza A Virus PA Antagonizes Interferon-beta by Interacting with Interferon Regulatory Factor 3. Front. Immunol. 2017 , 8 , 1051. [ Google Scholar ] [ CrossRef ]
- Wang, X.; Li, M.; Zheng, H.; Muster, T.; Palese, P.; Beg, A.A.; Garcia-Sastre, A. Influenza A virus NS1 protein prevents activation of NF-kappaB and induction of alpha/beta interferon. J. Virol. 2000 , 74 , 11566–11573. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Gao, S.; Song, L.; Li, J.; Zhang, Z.; Peng, H.; Jiang, W.; Wang, Q.; Kang, T.; Chen, S.; Huang, W. Influenza A virus-encoded NS1 virulence factor protein inhibits innate immune response by targeting IKK. Cell Microbiol. 2012 , 14 , 1849–1866. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Lee, M.C.; Yu, C.P.; Chen, X.H.; Liu, M.T.; Yang, J.R.; Chen, A.Y.; Huang, C.H. Influenza A virus NS1 protein represses antiviral immune response by hijacking NF-kappaB to mediate transcription of type III IFN. Front. Cell. Infect. Microbiol. 2022 , 12 , 998584. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Jagger, B.W.; Wise, H.M.; Kash, J.C.; Walters, K.A.; Wills, N.M.; Xiao, Y.L.; Dunfee, R.L.; Schwartzman, L.M.; Ozinsky, A.; Bell, G.L.; et al. An overlapping protein-coding region in influenza A virus segment 3 modulates the host response. Science 2012 , 337 , 199–204. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Hu, J.; Kong, M.; Cui, Z.; Gao, Z.; Ma, C.; Hu, Z.; Jiao, X.; Liu, X. PA-X protein of H5N1 avian influenza virus inhibits NF-kappaB activity, a potential mechanism for PA-X counteracting the host innate immune responses. Vet. Microbiol. 2020 , 250 , 108838. [ Google Scholar ] [ CrossRef ]
- Feng, M.; Zhang, Q.; Wu, W.; Chen, L.; Gu, S.; Ye, Y.; Zhong, Y.; Huang, Q.; Liu, S. Inducible Guanylate-Binding Protein 7 Facilitates Influenza A Virus Replication by Suppressing Innate Immunity via NF-kappaB and JAK-STAT Signaling Pathways. J. Virol. 2021 , 95 , JVI.02038-20. [ Google Scholar ] [ CrossRef ]
- Stasakova, J.; Ferko, B.; Kittel, C.; Sereinig, S.; Romanova, J.; Katinger, H.; Egorov, A. Influenza A mutant viruses with altered NS1 protein function provoke caspase-1 activation in primary human macrophages, resulting in fast apoptosis and release of high levels of interleukins 1beta and 18. J. Gen. Virol. 2005 , 86 , 185–195. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Moriyama, M.; Chen, I.Y.; Kawaguchi, A.; Koshiba, T.; Nagata, K.; Takeyama, H.; Hasegawa, H.; Ichinohe, T. The RNA- and TRIM25-Binding Domains of Influenza Virus NS1 Protein Are Essential for Suppression of NLRP3 Inflammasome-Mediated Interleukin-1beta Secretion. J. Virol. 2016 , 90 , 4105–4114. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Tao, P.; Ning, Z.; Zhou, P.; Xiao, W.; Wang, G.; Li, S.; Zhang, G. H3N2 canine influenza virus NS1 protein inhibits canine NLRP3 inflammasome activation. Vet. Immunol. Immunopathol. 2022 , 252 , 110483. [ Google Scholar ] [ CrossRef ]
- Chung, W.C.; Kang, H.R.; Yoon, H.; Kang, S.J.; Ting, J.P.; Song, M.J. Influenza A Virus NS1 Protein Inhibits the NLRP3 Inflammasome. PLoS ONE 2015 , 10 , e0126456. [ Google Scholar ] [ CrossRef ]
- Park, H.S.; Liu, G.; Thulasi Raman, S.N.; Landreth, S.L.; Liu, Q.; Zhou, Y. NS1 Protein of 2009 Pandemic Influenza A Virus Inhibits Porcine NLRP3 Inflammasome-Mediated Interleukin-1 Beta Production by Suppressing ASC Ubiquitination. J. Virol. 2018 , 92 , JVI.00022-18. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Boal-Carvalho, I.; Mazel-Sanchez, B.; Silva, F.; Garnier, L.; Yildiz, S.; Bonifacio, J.P.; Niu, C.; Williams, N.; Francois, P.; Schwerk, N.; et al. Influenza A viruses limit NLRP3-NEK7-complex formation and pyroptosis in human macrophages. EMBO Rep. 2020 , 21 , e50421. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Cheung, P.H.; Ye, Z.W.; Lee, T.T.; Chen, H.; Chan, C.P.; Jin, D.Y. PB1-F2 protein of highly pathogenic influenza A (H7N9) virus selectively suppresses RNA-induced NLRP3 inflammasome activation through inhibition of MAVS-NLRP3 interaction. J. Leukoc. Biol. 2020 , 108 , 1655–1663. [ Google Scholar ] [ CrossRef ]
- Silva, F.; Boal-Carvalho, I.; Williams, N.; Chabert, M.; Niu, C.; Hedhili, D.; Choltus, H.; Liaudet, N.; Gaia, N.; Karenovics, W.; et al. Identification of a short sequence motif in the influenza A virus pathogenicity factor PB1-F2 required for inhibition of human NLRP3. J. Virol. 2024 , 98 , e0041124. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Jia, D.; Rahbar, R.; Chan, R.W.; Lee, S.M.; Chan, M.C.; Wang, B.X.; Baker, D.P.; Sun, B.; Peiris, J.S.; Nicholls, J.M.; et al. Influenza virus non-structural protein 1 (NS1) disrupts interferon signaling. PLoS ONE 2010 , 5 , e13927. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Pauli, E.K.; Schmolke, M.; Wolff, T.; Viemann, D.; Roth, J.; Bode, J.G.; Ludwig, S. Influenza A virus inhibits type I IFN signaling via NF-kappaB-dependent induction of SOCS-3 expression. PLoS Pathog. 2008 , 4 , e1000196. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Du, Y.; Yang, F.; Wang, Q.; Xu, N.; Xie, Y.; Chen, S.; Qin, T.; Peng, D. Influenza a virus antagonizes type I and type II interferon responses via SOCS1-dependent ubiquitination and degradation of JAK1. Virol. J. 2020 , 17 , 74. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Yang, H.; Dong, Y.; Bian, Y.; Xu, N.; Wu, Y.; Yang, F.; Du, Y.; Qin, T.; Chen, S.; Peng, D.; et al. The influenza virus PB2 protein evades antiviral innate immunity by inhibiting JAK1/STAT signalling. Nat. Commun. 2022 , 13 , 6288. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Xia, C.; Vijayan, M.; Pritzl, C.J.; Fuchs, S.Y.; McDermott, A.B.; Hahm, B. Hemagglutinin of Influenza A Virus Antagonizes Type I Interferon (IFN) Responses by Inducing Degradation of Type I IFN Receptor 1. J. Virol. 2015 , 90 , 2403–2417. [ Google Scholar ] [ CrossRef ]
- Xia, C.; Wolf, J.J.; Vijayan, M.; Studstill, C.J.; Ma, W.; Hahm, B. Casein Kinase 1alpha Mediates the Degradation of Receptors for Type I and Type II Interferons Caused by Hemagglutinin of Influenza A Virus. J. Virol. 2018 , 92 , JVI.00006-18. [ Google Scholar ] [ CrossRef ]
- Lin, X.; Yu, S.; Ren, P.; Sun, X.; Jin, M. Human microRNA-30 inhibits influenza virus infection by suppressing the expression of SOCS1, SOCS3, and NEDD4. Cell Microbiol. 2020 , 22 , e13150. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Guo, M.; Li, F.; Ji, J.; Liu, Y.; Liu, F.; Zhao, Y.; Li, J.; Han, S.; Wang, Q.; Ding, G. Inhibition of miR-93 promotes interferon effector signaling to suppress influenza A infection by upregulating JAK1. Int. Immunopharmacol. 2020 , 86 , 106754. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Othumpangat, S.; Beezhold, D.H.; Umbright, C.M.; Noti, J.D. Influenza Virus-Induced Novel miRNAs Regulate the STAT Pathway. Viruses 2021 , 13 , 967. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Katze, M.G.; Tomita, J.; Black, T.; Krug, R.M.; Safer, B.; Hovanessian, A. Influenza virus regulates protein synthesis during infection by repressing autophosphorylation and activity of the cellular 68,000-Mr protein kinase. J. Virol. 1988 , 62 , 3710–3717. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Lu, Y.; Wambach, M.; Katze, M.G.; Krug, R.M. Binding of the influenza virus NS1 protein to double-stranded RNA inhibits the activation of the protein kinase that phosphorylates the elF-2 translation initiation factor. Virology 1995 , 214 , 222–228. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Bergmann, M.; Garcia-Sastre, A.; Carnero, E.; Pehamberger, H.; Wolff, K.; Palese, P.; Muster, T. Influenza virus NS1 protein counteracts PKR-mediated inhibition of replication. J. Virol. 2000 , 74 , 6203–6206. [ Google Scholar ] [ CrossRef ]
- Min, J.Y.; Krug, R.M. The primary function of RNA binding by the influenza A virus NS1 protein in infected cells: Inhibiting the 2′-5′ oligo (A) synthetase/RNase L pathway. Proc. Natl. Acad. Sci. USA 2006 , 103 , 7100–7105. [ Google Scholar ] [ CrossRef ]
- Zhu, Z.; Shi, Z.; Yan, W.; Wei, J.; Shao, D.; Deng, X.; Wang, S.; Li, B.; Tong, G.; Ma, Z. Nonstructural protein 1 of influenza A virus interacts with human guanylate-binding protein 1 to antagonize antiviral activity. PLoS ONE 2013 , 8 , e55920. [ Google Scholar ] [ CrossRef ]
- Hu, S.; Yin, L.; Mei, S.; Li, J.; Xu, F.; Sun, H.; Liu, X.; Cen, S.; Liang, C.; Li, A.; et al. BST-2 restricts IAV release and is countered by the viral M2 protein. Biochem. J. 2017 , 474 , 715–730. [ Google Scholar ] [ CrossRef ]
- Chesarino, N.M.; McMichael, T.M.; Yount, J.S. E3 Ubiquitin Ligase NEDD4 Promotes Influenza Virus Infection by Decreasing Levels of the Antiviral Protein IFITM3. PLoS Pathog. 2015 , 11 , e1005095. [ Google Scholar ] [ CrossRef ]
- Shan, J.; Zhao, B.; Shan, Z.; Nie, J.; Deng, R.; Xiong, R.; Tsun, A.; Pan, W.; Zhao, H.; Chen, L.; et al. Histone demethylase LSD1 restricts influenza A virus infection by erasing IFITM3-K88 monomethylation. PLoS Pathog. 2017 , 13 , e1006773. [ Google Scholar ] [ CrossRef ]
- Esposito, S.; Molteni, C.G.; Giliani, S.; Mazza, C.; Scala, A.; Tagliaferri, L.; Pelucchi, C.; Fossali, E.; Plebani, A.; Principi, N. Toll-like receptor 3 gene polymorphisms and severity of pandemic A/H1N1/2009 influenza in otherwise healthy children. Virol. J. 2012 , 9 , 270. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Lee, N.; Cao, B.; Ke, C.; Lu, H.; Hu, Y.; Tam, C.H.T.; Ma, R.C.W.; Guan, D.; Zhu, Z.; Li, H.; et al. IFITM3, TLR3, and CD55 Gene SNPs and Cumulative Genetic Risks for Severe Outcomes in Chinese Patients With H7N9/H1N1pdm09 Influenza. J. Infect. Dis. 2017 , 216 , 97–104. [ Google Scholar ] [ CrossRef ]
- Lim, H.K.; Huang, S.X.L.; Chen, J.; Kerner, G.; Gilliaux, O.; Bastard, P.; Dobbs, K.; Hernandez, N.; Goudin, N.; Hasek, M.L.; et al. Severe influenza pneumonitis in children with inherited TLR3 deficiency. J. Exp. Med. 2019 , 216 , 2038–2056. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Pryimenko, N.O.; Kotelevska, T.M.; Koval, T.I.; Syzova, L.M.; Dubynska, H.M.; Kaidashev Icapital Er, C. Genetic polymorphism ARG753GLN of TLR-2, LEU412PHE of TLR-3, ASP299GLY of TLR-4 in patients with influenza and influenza-associated pneumonia. Wiad. Lek. 2019 , 72 , 2324–2328. [ Google Scholar ] [ PubMed ]
- Bucciol, G.; Desmet, L.; Leuven Laboratory of Inborn Errors of Immunity; Corveleyn, A.; Meyts, I. Pathogenic P554S Variant in TLR3 in a Patient with Severe Influenza Pneumonia. J. Clin. Immunol. 2022 , 42 , 430–432. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Choudhary, M.L.; Chaudhary, U.; Salve, M.; Shinde, P.; Padbidri, V.; Sangle, S.A.; Salvi, S.; Bavdekar, A.R.; D’Costa, P.; Alagarasu, K. Functional Single-Nucleotide Polymorphisms in the MBL2 and TLR3 Genes Influence Disease Severity in Influenza A (H1N1)pdm09 Virus-Infected Patients from Maharashtra, India. Viral Immunol. 2022 , 35 , 303–309. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Jorgensen, S.E.; Christiansen, M.; Ryo, L.B.; Gad, H.H.; Gjedsted, J.; Staeheli, P.; Mikkelsen, J.G.; Storgaard, M.; Hartmann, R.; Mogensen, T.H. Defective RNA sensing by RIG-I in severe influenza virus infection. Clin. Exp. Immunol. 2018 , 192 , 366–376. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Lee, S.; Zhang, Y.; Newhams, M.; Novak, T.; Thomas, P.G.; Mourani, P.M.; Hall, M.W.; Loftis, L.L.; Cvijanovich, N.Z.; Tarquinio, K.M.; et al. DDX58 Is Associated With Susceptibility to Severe Influenza Virus Infection in Children and Adolescents. J. Infect. Dis. 2022 , 226 , 2030–2036. [ Google Scholar ] [ CrossRef ]
- Lee, M.N.; Ye, C.; Villani, A.C.; Raj, T.; Li, W.; Eisenhaure, T.M.; Imboywa, S.H.; Chipendo, P.I.; Ran, F.A.; Slowikowski, K.; et al. Common genetic variants modulate pathogen-sensing responses in human dendritic cells. Science 2014 , 343 , 1246980. [ Google Scholar ] [ CrossRef ]
- Ciancanelli, M.J.; Huang, S.X.; Luthra, P.; Garner, H.; Itan, Y.; Volpi, S.; Lafaille, F.G.; Trouillet, C.; Schmolke, M.; Albrecht, R.A.; et al. Infectious disease. Life-threatening influenza and impaired interferon amplification in human IRF7 deficiency. Science 2015 , 348 , 448–453. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Hernandez, N.; Melki, I.; Jing, H.; Habib, T.; Huang, S.S.Y.; Danielson, J.; Kula, T.; Drutman, S.; Belkaya, S.; Rattina, V.; et al. Life-threatening influenza pneumonitis in a child with inherited IRF9 deficiency. J. Exp. Med. 2018 , 215 , 2567–2585. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Thomsen, M.M.; Jorgensen, S.E.; Gad, H.H.; Storgaard, M.; Gjedsted, J.; Christiansen, M.; Hartmann, R.; Mogensen, T.H. Defective interferon priming and impaired antiviral responses in a patient with an IRF7 variant and severe influenza. Med. Microbiol. Immunol. 2019 , 208 , 869–876. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Calik Basaran, N.; Tan, C.; Ozisik, L.; Ozbek, B.; Inkaya, A.C.; Alp, S.; Ersoy, E.O.; Ayvaz, D.C.; Tezcan, I. Evaluation of the peripheral blood T and B cell subsets and IRF-7 variants in adult patients with severe influenza virus infection. Health Sci. Rep. 2022 , 5 , e492. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Chen, Y.; Graf, L.; Chen, T.; Liao, Q.; Bai, T.; Petric, P.P.; Zhu, W.; Yang, L.; Dong, J.; Lu, J.; et al. Rare variant MX1 alleles increase human susceptibility to zoonotic H7N9 influenza virus. Science 2021 , 373 , 918–922. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Graf, L.; Dick, A.; Sendker, F.; Barth, E.; Marz, M.; Daumke, O.; Kochs, G. Effects of allelic variations in the human myxovirus resistance protein A on its antiviral activity. J. Biol. Chem. 2018 , 293 , 3056–3072. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Mitchell, P.S.; Patzina, C.; Emerman, M.; Haller, O.; Malik, H.S.; Kochs, G. Evolution-guided identification of antiviral specificity determinants in the broadly acting interferon-induced innate immunity factor MxA. Cell Host Microbe 2012 , 12 , 598–604. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Dittmann, J.; Stertz, S.; Grimm, D.; Steel, J.; Garcia-Sastre, A.; Haller, O.; Kochs, G. Influenza A virus strains differ in sensitivity to the antiviral action of Mx-GTPase. J. Virol. 2008 , 82 , 3624–3631. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Zimmermann, P.; Manz, B.; Haller, O.; Schwemmle, M.; Kochs, G. The viral nucleoprotein determines Mx sensitivity of influenza A viruses. J. Virol. 2011 , 85 , 8133–8140. [ Google Scholar ] [ CrossRef ]
- Manz, B.; Dornfeld, D.; Gotz, V.; Zell, R.; Zimmermann, P.; Haller, O.; Kochs, G.; Schwemmle, M. Pandemic influenza A viruses escape from restriction by human MxA through adaptive mutations in the nucleoprotein. PLoS Pathog. 2013 , 9 , e1003279. [ Google Scholar ] [ CrossRef ]
- Riegger, D.; Hai, R.; Dornfeld, D.; Manz, B.; Leyva-Grado, V.; Sanchez-Aparicio, M.T.; Albrecht, R.A.; Palese, P.; Haller, O.; Schwemmle, M.; et al. The nucleoprotein of newly emerged H7N9 influenza A virus harbors a unique motif conferring resistance to antiviral human MxA. J. Virol. 2015 , 89 , 2241–2252. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Gotz, V.; Magar, L.; Dornfeld, D.; Giese, S.; Pohlmann, A.; Hoper, D.; Kong, B.W.; Jans, D.A.; Beer, M.; Haller, O.; et al. Influenza A viruses escape from MxA restriction at the expense of efficient nuclear vRNP import. Sci. Rep. 2016 , 6 , 23138. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Deeg, C.M.; Hassan, E.; Mutz, P.; Rheinemann, L.; Gotz, V.; Magar, L.; Schilling, M.; Kallfass, C.; Nurnberger, C.; Soubies, S.; et al. In vivo evasion of MxA by avian influenza viruses requires human signature in the viral nucleoprotein. J. Exp. Med. 2017 , 214 , 1239–1248. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Dornfeld, D.; Petric, P.P.; Hassan, E.; Zell, R.; Schwemmle, M. Eurasian Avian-Like Swine Influenza A Viruses Escape Human MxA Restriction through Distinct Mutations in Their Nucleoprotein. J. Virol. 2019 , 93 , JVI.00997-00918. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Everitt, A.R.; Clare, S.; Pertel, T.; John, S.P.; Wash, R.S.; Smith, S.E.; Chin, C.R.; Feeley, E.M.; Sims, J.S.; Adams, D.J.; et al. IFITM3 restricts the morbidity and mortality associated with influenza. Nature 2012 , 484 , 519–523. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Allen, E.K.; Randolph, A.G.; Bhangale, T.; Dogra, P.; Ohlson, M.; Oshansky, C.M.; Zamora, A.E.; Shannon, J.P.; Finkelstein, D.; Dressen, A.; et al. SNP-mediated disruption of CTCF binding at the IFITM3 promoter is associated with risk of severe influenza in humans. Nat. Med. 2017 , 23 , 975–983. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Zhang, Y.H.; Zhao, Y.; Li, N.; Peng, Y.C.; Giannoulatou, E.; Jin, R.H.; Yan, H.P.; Wu, H.; Liu, J.H.; Liu, N.; et al. Interferon-induced transmembrane protein-3 genetic variant rs12252-C is associated with severe influenza in Chinese individuals. Nat. Commun. 2013 , 4 , 1418. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Pan, Y.; Yang, P.; Dong, T.; Zhang, Y.; Shi, W.; Peng, X.; Cui, S.; Zhang, D.; Lu, G.; Liu, Y.; et al. IFITM3 Rs12252-C Variant Increases Potential Risk for Severe Influenza Virus Infection in Chinese Population. Front. Cell. Infect. Microbiol. 2017 , 7 , 294. [ Google Scholar ] [ CrossRef ]
- Kim, Y.C.; Jeong, M.J.; Jeong, B.H. Strong association of regulatory single nucleotide polymorphisms (SNPs) of the IFITM3 gene with influenza H1N1 2009 pandemic virus infection. Cell Mol. Immunol. 2020 , 17 , 662–664. [ Google Scholar ] [ CrossRef ]
- Wang, Z.; Zhang, A.; Wan, Y.; Liu, X.; Qiu, C.; Xi, X.; Ren, Y.; Wang, J.; Dong, Y.; Bao, M.; et al. Early hypercytokinemia is associated with interferon-induced transmembrane protein-3 dysfunction and predictive of fatal H7N9 infection. Proc. Natl. Acad. Sci. USA 2014 , 111 , 769–774. [ Google Scholar ] [ CrossRef ]
- Xuan, Y.; Wang, L.N.; Li, W.; Zi, H.R.; Guo, Y.; Yan, W.J.; Chen, X.B.; Wei, P.M. IFITM3 rs12252 T>C polymorphism is associated with the risk of severe influenza: A meta-analysis. Epidemiol. Infect. 2015 , 143 , 2975–2984. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Kim, Y.C.; Jeong, B.H. Ethnic variation in risk genotypes based on single nucleotide polymorphisms (SNPs) of the interferon-inducible transmembrane 3 (IFITM3) gene, a susceptibility factor for pandemic 2009 H1N1 influenza A virus. Immunogenetics 2020 , 72 , 447–453. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Yang, X.; Tan, B.; Zhou, X.; Xue, J.; Zhang, X.; Wang, P.; Shao, C.; Li, Y.; Li, C.; Xia, H.; et al. Interferon-Inducible Transmembrane Protein 3 Genetic Variant rs12252 and Influenza Susceptibility and Severity: A Meta-Analysis. PLoS ONE 2015 , 10 , e0124985. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Mehrbod, P.; Eybpoosh, S.; Fotouhi, F.; Shokouhi Targhi, H.; Mazaheri, V.; Farahmand, B. Association of IFITM3 rs12252 polymorphisms, BMI, diabetes, and hypercholesterolemia with mild flu in an Iranian population. Virol. J. 2017 , 14 , 218. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Jia, R.; Pan, Q.; Ding, S.; Rong, L.; Liu, S.L.; Geng, Y.; Qiao, W.; Liang, C. The N-terminal region of IFITM3 modulates its antiviral activity by regulating IFITM3 cellular localization. J. Virol. 2012 , 86 , 13697–13707. [ Google Scholar ] [ CrossRef ]
- Jia, R.; Xu, F.; Qian, J.; Yao, Y.; Miao, C.; Zheng, Y.M.; Liu, S.L.; Guo, F.; Geng, Y.; Qiao, W.; et al. Identification of an endocytic signal essential for the antiviral action of IFITM3. Cell Microbiol. 2014 , 16 , 1080–1093. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- John, S.P.; Chin, C.R.; Perreira, J.M.; Feeley, E.M.; Aker, A.M.; Savidis, G.; Smith, S.E.; Elia, A.E.; Everitt, A.R.; Vora, M.; et al. The CD225 domain of IFITM3 is required for both IFITM protein association and inhibition of influenza A virus and dengue virus replication. J. Virol. 2013 , 87 , 7837–7852. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Hensen, L.; Matrosovich, T.; Roth, K.; Klenk, H.D.; Matrosovich, M. HA-Dependent Tropism of H5N1 and H7N9 Influenza Viruses to Human Endothelial Cells Is Determined by Reduced Stability of the HA, Which Allows the Virus To Cope with Inefficient Endosomal Acidification and Constitutively Expressed IFITM3. J. Virol. 2019 , 94 , JVI.01223-19. [ Google Scholar ] [ CrossRef ]
- Sanchez-Gonzalez, M.T.; Cienfuegos-Jimenez, O.; Alvarez-Cuevas, S.; Perez-Maya, A.A.; Borrego-Soto, G.; Marino-Martinez, I.A. Prevalence of the SNP rs10774671 of the OAS1 gene in Mexico as a possible predisposing factor for RNA virus disease. Int. J. Mol. Epidemiol. Genet. 2021 , 12 , 52–60. [ Google Scholar ]
- Dittmann, M.; Hoffmann, H.H.; Scull, M.A.; Gilmore, R.H.; Bell, K.L.; Ciancanelli, M.; Wilson, S.J.; Crotta, S.; Yu, Y.; Flatley, B.; et al. A serpin shapes the extracellular environment to prevent influenza A virus maturation. Cell 2015 , 160 , 631–643. [ Google Scholar ] [ CrossRef ]
- World Health Organization, WHO. Seasonal Influenza Factsheet. Available online: https://www.who.int/news-room/fact-sheets/detail/influenza-(seasonal) (accessed on 10 January 2024).
- Caserta, L.C.; Frye, E.A.; Butt, S.L.; Laverack, M.A.; Nooruzzaman, M.; Covalenda, L.M.; Thompson, A.; Prarat Koscielny, M.; Cronk, B.; Johnson, A.; et al. From birds to mammals: Spillover of highly pathogenic avian influenza H5N1 virus to dairy cattle led to efficient intra- and interspecies transmission. bioRxiv 2024 . [ Google Scholar ] [ CrossRef ]
- Ly, H. Highly pathogenic avian influenza H5N1 virus infections of dairy cattle and livestock handlers in the United States of America. Virulence 2024 , 15 , 2343931. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Uyeki, T.M.; Milton, S.; Abdul Hamid, C.; Reinoso Webb, C.; Presley, S.M.; Shetty, V.; Rollo, S.N.; Martinez, D.L.; Rai, S.; Gonzales, E.R.; et al. Highly Pathogenic Avian Influenza A(H5N1) Virus Infection in a Dairy Farm Worker. N. Engl. J. Med. 2024 , 390 , 2028–2029. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Swierczynska, M.; Mirowska-Guzel, D.M.; Pindelska, E. Antiviral Drugs in Influenza. Int. J. Environ. Res. Public Health 2022 , 19 , 3018. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Kumari, R.; Sharma, S.D.; Kumar, A.; Ende, Z.; Mishina, M.; Wang, Y.; Falls, Z.; Samudrala, R.; Pohl, J.; Knight, P.R.; et al. Antiviral Approaches against Influenza Virus. Clin. Microbiol. Rev. 2023 , 36 , e0004022. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Hussain, M.; Galvin, H.D.; Haw, T.Y.; Nutsford, A.N.; Husain, M. Drug resistance in influenza A virus: The epidemiology and management. Infect. Drug Resist. 2017 , 10 , 121–134. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Kumar, N.; Sharma, S.; Kumar, R.; Tripathi, B.N.; Barua, S.; Ly, H.; Rouse, B.T. Host-Directed Antiviral Therapy. Clin. Microbiol. Rev. 2020 , 33 , 00168-19. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Trimarco, J.D.; Heaton, N.S. From high-throughput to therapeutic: Host-directed interventions against influenza viruses. Curr. Opin. Virol. 2022 , 53 , 101198. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Girkin, J.L.N.; Maltby, S.; Bartlett, N.W. Toll-like receptor-agonist-based therapies for respiratory viral diseases: Thinking outside the cell. Eur. Respir. Rev. 2022 , 31 , 210274. [ Google Scholar ] [ CrossRef ]
- Kayesh, M.E.H.; Kohara, M.; Tsukiyama-Kohara, K. TLR agonists as vaccine adjuvants in the prevention of viral infections: An overview. Front. Microbiol. 2023 , 14 , 1249718. [ Google Scholar ] [ CrossRef ]
- Gautam, A.; Boyd, D.F.; Nikhar, S.; Zhang, T.; Siokas, I.; Van de Velde, L.A.; Gaevert, J.; Meliopoulos, V.; Thapa, B.; Rodriguez, D.A.; et al. Necroptosis blockade prevents lung injury in severe influenza. Nature 2024 , 628 , 835–843. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Click here to enlarge figure
| Genes | Variants/Mutations | Effect |
|---|---|---|
| TLR3 | rs5743313/CT, rs5743313, L412F, P554S/P680L, rs3775291/rs3775290 | Escape from TLR3-mediated signaling |
| RIG-I (DDX58) | p.R71H/p.P885S, rs4487862 | Escape from RIG-I-mediated signaling |
| IRF7 | p.Phe410Val (F410V)/p.Gln421X(Q421X), E331V | Escape from weakened IFN response |
| IRF9 | c.991G > A | Escape from weakened IFN response |
| MxA | Mutations in GTPase domain | Escape from MxA-mediated restriction |
| IFITM3 | rs12252-C | Escape from IFITM3-mediated restriction |
| OAS | rs10774671 | Escape from OAS-mediated restriction |
| Serpin 1 | rs6092 | Escape from Serpin 1-mediated restriction |
| The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
Share and Cite
An, W.; Lakhina, S.; Leong, J.; Rawat, K.; Husain, M. Host Innate Antiviral Response to Influenza A Virus Infection: From Viral Sensing to Antagonism and Escape. Pathogens 2024 , 13 , 561. https://doi.org/10.3390/pathogens13070561
An W, Lakhina S, Leong J, Rawat K, Husain M. Host Innate Antiviral Response to Influenza A Virus Infection: From Viral Sensing to Antagonism and Escape. Pathogens . 2024; 13(7):561. https://doi.org/10.3390/pathogens13070561
An, Wenlong, Simran Lakhina, Jessica Leong, Kartik Rawat, and Matloob Husain. 2024. "Host Innate Antiviral Response to Influenza A Virus Infection: From Viral Sensing to Antagonism and Escape" Pathogens 13, no. 7: 561. https://doi.org/10.3390/pathogens13070561
Article Metrics
Article access statistics, further information, mdpi initiatives, follow mdpi.
Subscribe to receive issue release notifications and newsletters from MDPI journals
A .gov website belongs to an official government organization in the United States.
A lock ( ) or https:// means you've safely connected to the .gov website. Share sensitive information only on official, secure websites.
- About Respiratory Illnesses
- Risk Factors
- Data and Trends
- Resources to Prepare for Flu, COVID-19, and RSV
- Respiratory Virus Guidance
- Health Care Provider Toolkit
- Long-Term Care Toolkit
Related Topics:
- View All Home
- Seasonal Flu
Long-Term Care Quick Start Guide: Preparing for Respiratory Virus Season
What to know.
When respiratory viruses are spreading in the community, the risk for spread in long-term care (LTC) settings, including nursing homes, increases as well. Existing infection prevention and control measures can help prevent and slow the spread of flu, RSV, and COVID-19 to protect residents and staff. Preparing and responding to respiratory viruses in LTC settings requires a comprehensive approach that includes infection prevention, testing, vaccination, and treatment.
Infection Prevention and Testing
Implement recommended infection prevention and control practices in the facility. Be aware when respiratory virus levels are increasing in the community and ensure access to respiratory viral testing with rapid results.
Guidance and recommendations
- Preventing Transmission of Viral Respiratory Pathogens in Healthcare Settings
- Viral Respiratory Pathogens Toolkit for Nursing Homes
- Testing and Management Considerations for Nursing Home Residents with Acute Respiratory Illness Symptoms When SARS-CoV-2 and Influenza Viruses Are Co-circulating
- Infection Control: Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2)
- Interim Guidance for Influenza Outbreak Management in Long-Term Care and Post-Acute Care Facilities
- Poster: Infection Control Actions for Respiratory Viruses
- Masking Signage for LTC: Wear A Mask Illustration and Wear A Mask Photo
- Project Firstline: Infection Control Actions for Respiratory Viruses
Vaccination
Provide recommended flu, RSV, and COVID-19 vaccines to residents and staff and encourage visitors to be vaccinated.
Guidance and Recommendations
- ACIP Seasonal Influenza Vaccine Recommendations
- ACIP Influenza Summary 2023-2024
- ACIP RSV Vaccine Recommendations
- Healthcare Providers: RSV Vaccination for Adults 60 Years of Age and Over
- ACIP COVID-19 Vaccine Recommendations
- Clinical Guidance for COVID-19 Vaccination
Resources for Providers
- Immunization Schedules: Immunization Schedules for Healthcare Providers
- Toolkit: Preparing Your Patients for the Fall and Winter Virus Season
- Flu: Seasonal Influenza Vaccination Resources for Health Professionals
- COVID-19: Vaccination Clinical & Professional Resources
- Shared Clinical Decision-Making Handout: RSV Vaccination for Adults 60 Years and Older
- Fact Sheet: RSV Fact Sheet for Healthcare Providers
- Frequently Asked Questions: 2023-2024 Vaccine Recommendations for Long-term Care Facilities
Resources for Patients and Families
- Flu: Flu & People 65 Years and Older
- RSV: RSV Vaccination for Older Adults 60 Years of Age and Over
- COVID-19 Vaccines: COVID-19 Vaccines for Long-term Care Residents
- Flu and COVID-19 Vaccine Coadministration: Getting a Flu Vaccine and a COVID-19 Vaccine at the Same Time
- Fact Sheet: People 65 Years and Older Need a Flu Shot
- Fact Sheet: RSV in Older Adults
Provide residents timely access to treatment and/or prophylaxis for flu and COVID-19.
- Influenza Antiviral Medications: Summary for Clinicians
- Interim Clinical Considerations for COVID-19 Treatment in Outpatients
- Poster: What You Should Know About Influenza (Flu) Antiviral Drugs
- Job Aid: COVID-19 Therapeutics Decision Aid
- Chart: Side-by-Side Overview of Therapeutics Authorized or Approved for the Prevention of COVID-19 Infection or Treatment of Mild to Moderate COVID-19
Reporting to the National Healthcare Safety Network
Data nursing homes report to CDC's National Healthcare Safety Network (NHSN) are critical to understanding the impact of COVID-19 and COVID-19 vaccination in older adults, as well as supporting efforts to prevent infections.
- Long Term Care HCP & Resident COVID-19 Vaccination
- COVID-19 Module
Respiratory Illnesses
Respiratory illnesses affect the lungs and airways. Get information on common causes, prevention actions, and current levels of illness in your community.
For Everyone
Health care providers.
- Remote Access
- Save figures into PowerPoint
- Download tables as PDFs

Chapter 13: Viral Infections of the Skin
Erin Luxenberg
- Download Chapter PDF
Disclaimer: These citations have been automatically generated based on the information we have and it may not be 100% accurate. Please consult the latest official manual style if you have any questions regarding the format accuracy.
Download citation file:
- Search Book
Jump to a Section
Introduction to chapter, introduction, pathogenesis, clinical presentation.
- Clinical Course and Prognosis
- Indications for Consultation
- Patient Information
- Public Health Issues
- Differential Diagnosis
- Management of Nongenital Warts
- Management of Genital Warts
- Full Chapter
- Supplementary Content
Despite all the advances in antiviral therapy and the body's efficient immune system, the viruses that cause common skin infections continue to evade complete destruction. The herpes simplex and herpes zoster virus can persist in a dormant state in the dorsal root ganglia. The viruses that cause verrucae vulgaris (common warts) and molluscum contagiosum can persist for months to several years in the epidermis.
Herpes simplex and herpes zoster infections can cause significant illness and death especially in immunocompromised patients if the infection spreads to other organs. Common nongenital warts and molluscum contagiosum rarely cause significant problems in immunocompetent patients, but for various reasons most patients want treatment for these conditions. Genital warts are often asymptomatic and may be clinically undetectable; however, patients with oncogenic wart virus infections are at increased risk for anogenital and oropharyngeal cancers. 1
HERPES SIMPLEX
Table Key Points for Herpes Simplex
✓ Herpes simplex viruses (HSV-1 and HSV-2) are double - stranded DNA viruses that cause primary, latent and recurrent infections.
✓ HSV presents with painful grouped vesicles that often erode and develop crusts.
✓ Topical and oral HSV antiviral medications are most effective if given within 72 hours of the onset of symptoms.
Herpes simplex viruses (HSV) cause primary, latent, and recurrent infections and they are common infections worldwide. Human herpes virus-1 (HSV-1) typically infects the oral cavity, lips, and perioral skin and is usually acquired in childhood via nonsexual contact. Human herpes virus-2 (HSV-2) primarily infects the genital area and is almost always acquired via sexual contact. Nevertheless, HSV-1 is becoming a more common cause of genital herpes infections 1 . HSV has worldwide distribution but may be more common in less developed countries. Antibodies to HSV-1 are present in up to 85% of adults and antibodies to HSV-2 are present in 20–25% of adults. 2 However, many patients who have antibodies to HSV do not recall having had an infection.
HSV-1 and HSV-2 are human herpesviruses (HHV) which have double - stranded DNA and replicate within the nuclei of infected cells. HSV infects mucocutaneous tissue after direct contact or by way of secretions, mainly saliva in the case of HSV-1. The virus is transmitted via sensory nerves to the ganglia, where it may reside in a latent stage. Recurrent infections are caused by reactivation of the virus, which travels back to the skin or mucous membranes resulting in an active infection. Immune mechanisms suppress the virus with clearing of the lesions in 1–2 weeks, but latency in the ganglia persists. Recurrent mucocutaneous infection may occur every few weeks to months to years via viral reactivation. Viral shedding may continue after the infection has clinically resolved.
Get Free Access Through Your Institution
Pop-up div successfully displayed.
This div only appears when the trigger link is hovered over. Otherwise it is hidden from view.
Please Wait
Thank you for visiting nature.com. You are using a browser version with limited support for CSS. To obtain the best experience, we recommend you use a more up to date browser (or turn off compatibility mode in Internet Explorer). In the meantime, to ensure continued support, we are displaying the site without styles and JavaScript.
- View all journals
- Explore content
- About the journal
- Publish with us
- Sign up for alerts
- 12 July 2024
Bird flu could become a human pandemic. How are countries preparing?
- Smriti Mallapaty
You can also search for this author in PubMed Google Scholar
Highly pathogenic avian influenza has been detected in 145 cattle herds in the United States. Credit: Matthew Ludak for The Washington Post via Getty
As cases of avian influenza continue to rise in cattle in the United States, countries are preparing for the possibility that the virus could start spreading in people. Many nations are ramping up surveillance, as well as purchasing vaccines or developing new ones.
“This virus in its current state does not look like it has the characteristics of causing a pandemic. But with influenza viruses, that equation could entirely change with a single mutation,” says Scott Hensley, an immunologist at the University of Pennsylvania in Philadelphia.
The highly pathogenic avian influenza H5N1 has so far been detected in 145 cattle herds and 4 farm workers in a dozen states across the United States. Researchers say many more cases in cows and people have probably gone undetected . The chances of quashing the outbreak get “more slim by the day”, says Angela Rasmussen, a virologist at the University of Saskatchewan in Saskatoon, Canada.
Studies suggest that the virus is spreading between cows through contaminated milking equipment 1 , 2 , rather than airborne particles. The biggest risk is that it could evolve to infect mammals more effectively, including through the respiratory system, which would make it more difficult to contain. Given the close and regular contact that cows have with people, airborne transmission could spark a pandemic.
Efforts to prepare for that possibility include risk assessments, modelling and outbreak predictions. “There is loads of planning and preparedness going on internationally,” says Michelle Wille, a virus ecologist at the University of Melbourne in Australia.
Nicole Lurie, who heads preparedness and response at the Oslo-based Coalition for Epidemic Preparedness Innovations (CEPI), says the coalition’s approach “for the moment is one of ‘calm urgency’” — “like putting our shoes on in case we need to start running”.
Vaccinating people
A key focus of pandemic preparedness efforts is vaccines, which would protect people from getting ill should the virus spread more widely. Vaccinating people would also reduce the risk of H5N1 mixing with seasonal influenza viruses that are already well-adapted to spread in humans.
In May, the World Health Organization in Geneva, Switzerland, initiated a review of available influenza candidate vaccines, and confirmed that they would work against the H5N1 virus circulating in cattle. “Although the current public health risk is low, WHO is operating in a constant state of readiness for a potential influenza pandemic,” says Maria Van Kerkhove, who heads epidemic and pandemic preparedness and prevention at the WHO.
Last month, the European Commission purchased roughly 700,000 doses of a flu vaccine manufactured by CSL Seqirus, in Maidenhead, UK, with the option to buy another 40 million. The vaccine protects against H5 strains of influenza A. Also in June, Finland began vaccinating people against avian influenza, focusing on high-risk workers at fur and poultry farms.
Other countries, especially the United States, should also consider vaccinating high-risk workers, says Rasmussen. In May, the US Department of Health and Human Services (HHS) purchased almost five million more doses of the CSL Seqirus influenza vaccine for its stockpile.
But currently available vaccines rely on inactivated strains of viruses grown in chicken eggs, which are cheap, but slow, to produce. Researchers are developing vaccines using mRNA technology ; these are more expensive but quicker to manufacture, and their formulation can be updated to target emerging strains. “It really is a game-changer,” says Hensley, who has developed an H5 mRNA vaccine candidate and tested it in ferrets 3 . “In the case of a pandemic, you can expect that these vaccines will be used widely.”
Last week, the HHS announced that it had provided the pharmaceutical company Moderna, based in Cambridge, Massachusetts, with US$176 million to develop an mRNA-based vaccine against H5 influenza.
CEPI is working to ensure that the response is equitable worldwide. Half of existing vaccine supplies are already tied up in contracts or export controls, says Lurie, and it’s important to make sure that the remaining doses reach the people who need them. “As we saw during the COVID-19 pandemic, low- and middle-income countries could once again be pushed to the back of the queue.”
Doses for cows
Countries including the United States are investigating the possibility of vaccinating cattle to reduce transmission. “This could be a phenomenal mitigation effort” and would be practical to implement as part of existing drives to vaccinate livestock, says Jenna Guthmiller, an immunologist at the University of Colorado Anschutz Medical Campus in Aurora.
Several research teams are in the early stages of developing vaccines for cattle. But there are challenges to overcome. Studies suggest that the virus spreading in cattle finds safe harbour in the mammary glands and epithelial cells 2 of the udder. This could be a challenging site in which to elicit a protective immune response, says Diego Diel, a virologist at Cornell University in Ithaca, New York, who is developing candidate vaccines against highly pathogenic avian influenza that use harmless DNA viruses to deliver genetic material. Hensley is currently testing his mRNA vaccine in cattle and swine.
But one concern is that vaccines could cover up symptoms in animals that are still infectious, which would increase the risk to people, says Thomas Peacock, a virologist at Imperial College London.
Vaccines should be seen as a measure of last resort, after implementing all other layers of containment, says Martin Beer, a virologist at the Federal Research Institute for Animal Health in Greifswald, Germany. They protect against “a worst-case scenario”.
Surveillance
To stay ahead of the virus, countries are also tracking its spread through increased testing of people and animals. Before the US outbreak, researchers didn’t think cattle could be infected with avian influenza. They are now scrambling to develop tests specific to this host.
Isabella Monne, who studies the molecular epidemiology of animal viruses at the Experimental Zooprophylactic Institute of Venice in Legnaro, Italy, is developing and evaluating tools to help laboratories across Europe to detect viral particles and antibodies, which are evidence of past infection, in cow blood and milk. Groups across Europe, Canada and the United States have started testing cow blood or bulk milk samples.
Researchers are also monitoring sequences of the virus’s genome for changes that would improve its ability to infect cells found in the upper airways. These mutations would increase the risk to people.
One group has created 4 a library of every possible amino-acid mutation on the haemagglutinin protein, which the virus uses to enter cells. The researchers tested in human cells how well the mutated proteins bind to upper-airway receptors, and their stability in acidic environments — traits “known to correlate with viruses going from avian to mammalian hosts, and becoming pandemics”, says Peacock, a co-author of the study, which has not been peer reviewed. Scanning for those mutations could allow real-time risk prediction, he says.
doi: https://doi.org/10.1038/d41586-024-02237-4
Le Sage, V. et al. Preprint at bioRxiv https://doi.org/10.1101/2024.05.22.595317 (2024).
Caserta, L. C. et al. Preprint at bioRxiv https://doi.org/content/10.1101/2024.05.22.595317 (2024).
Furey, C. et al. Nature Commun. 15, 4350 (2024).
Article Google Scholar
Dadonaite, B. et al. Preprint at bioRxiv https://doi.org/10.1101/2024.05.23.595634 (2024).
Download references
Reprints and permissions
Related Articles

- Public health
- Epidemiology
The pandemic agreement: an African perspective
Correspondence 16 JUL 24

These period pads solidify blood to prevent leaks
News 10 JUL 24

If bird flu sparks a human pandemic, your past immunity could help
News 09 JUL 24

Vaccines save lives: how can uptake be increased?
Editorial 09 JUL 24

Combined COVID–flu vaccines are coming: Moderna jab clears major test
News 28 JUN 24

Hotspot shelters stimulate frog resistance to chytridiomycosis
Article 26 JUN 24

Repeated plague infections across six generations of Neolithic Farmers
Article 10 JUL 24

Could bird flu in cows lead to a human outbreak? Slow response worries scientists
News 17 MAY 24

Bird flu in US cows: where will it end?
News 08 MAY 24
Editor (Electrical and Electronic Engineering)
The Associate/Senior Editor role at Nature Communications is ideal for researchers who love science and engineering.
London, Berlin or Pune – hybrid working model.
Springer Nature Ltd
High-Level Talents at the First Affiliated Hospital of Nanchang University
For clinical medicine and basic medicine; basic research of emerging inter-disciplines and medical big data.
Nanchang, Jiangxi, China
The First Affiliated Hospital of Nanchang University
Associate or Senior Editor, Nature Electronics
Job Title: Associate or Senior Editor, Nature Electronics Location: New York, Shanghai or Beijing — Hybrid Working Closing date for application...
New York City, New York (US)
Postdoctoral Fellow
To study how a population of skeletal stem cells mediate bone regeneration and orchestrate a composite tissue regeneration program using mouse models.
Hospital for Special Surgery, New York City, New York (US)
Hospital for Special Surgery
Lecturer / Associate Professor in Advanced Cell Models
Lecturer / Associate Professor in Advanced Cell Models Cell and Developmental Biology Location: Highfield Campus Salary: £44,263 to £72,018 per ann...
Southampton, Hampshire (GB)
University of Southampton
Sign up for the Nature Briefing newsletter — what matters in science, free to your inbox daily.
Quick links
- Explore articles by subject
- Guide to authors
- Editorial policies
- U.S. Department of Health & Human Services
- Virtual Tour
- Staff Directory
- En Español
You are here
News releases.
Media Advisory
Monday, July 8, 2024
Features of H5N1 influenza viruses in dairy cows may facilitate infection, transmission in mammals
H5N1 virus did not efficiently transmit via respiratory route to ferrets.

A series of experiments with highly pathogenic H5N1 avian influenza (HPAI H5N1) viruses circulating in infected U.S. dairy cattle found that viruses derived from lactating dairy cattle induced severe disease in mice and ferrets when administered via intranasal inoculation. The virus from the H5N1-infected cows bound to both avian (bird) and human-type cellular receptors, but, importantly, did not transmit efficiently among ferrets exposed via respiratory droplets. The findings, published in Nature , suggest that bovine (cow) HPAI H5N1 viruses may differ from previous HPAI H5N1 viruses and that these viruses may possess features that could facilitate infection and transmission among mammals. However, they currently do not appear capable of efficient respiratory transmission between animals or people.
In March 2024, an outbreak of HPAI H5N1 was reported among U.S. dairy cattle which spread across herds and led to fatal infections among some cats on affected farms, spillover into poultry, and four reported infections among dairy workers. The HPAI H5N1 viruses isolated from affected cattle are closely related to H5N1 viruses that have circulated in North American wild birds since late 2021. Over time, those avian viruses have undergone genetic changes and have spread throughout the continent causing outbreaks in wild birds and mammals—sometimes with mortality rates and suspected transmission within species.
To better understand the characteristics of the bovine H5N1 viruses, researchers from the University of Wisconsin-Madison, Japan’s Shizuoka and Tokyo Universities, and Texas A&M Veterinary Medical Diagnostic Laboratory conducted experiments to determine the ability of bovine HPAI H5N1 to replicate and cause disease in mice and ferrets, which are routinely used for influenza A virus studies. Ferrets are thought to be a good model for understanding potential influenza transmission patterns in people because they exhibit similar clinical symptoms, immune responses and develop respiratory tract infections like humans.
The researchers intranasally administered to mice doses of bovine HPAI H5N1 influenza of increasing strength (5 mice per dosage group), and then monitored the animals for body weight changes and survival for 15 days. All the mice that received the higher doses died of infection. Some of the mice that received lower doses survived, and those that received the lowest dose experienced no body weight loss and survived.
The researchers also compared the effects of the bovine HPAI H5N1 virus to a Vietnamese H5N1 strain that is typical of H5N1 avian influenza virus in humans and to an H1N1 influenza virus, both delivered intranasally to mice. The mice that received either the bovine HPAI H5N1 virus or the Vietnamese avian H5N1 virus experienced high virus levels in respiratory and non-respiratory organs, including in the mammary glands and muscle tissues, and sporadic detection in the eyes. The H1N1 virus was found only in the respiratory tissues of the animals. Ferrets intranasally infected with the bovine HPAI H5N1 virus experienced elevated temperatures and loss of body weight. As with the mice, the scientists discovered high virus levels in the ferrets’ upper and lower respiratory tracts and other organs. Unlike the mice, however, no virus was found in the ferrets’ blood or muscle tissues.
“Together, our pathogenicity studies in mice and ferrets revealed that HPAI H5N1 derived from lactating dairy cattle may induce severe disease after oral ingestion or respiratory infection, and infection by either the oral or respiratory route can lead to systemic spread of virus to non-respiratory tissues including the eye, mammary gland, teat and/or muscle,” the authors write.
To test whether bovine H5N1 viruses transmit among mammals via respiratory droplets, such as emitted by coughs and sneezes, the researchers infected groups of ferrets (four animals per group) with either bovine HPAI H5N1 virus or H1N1 influenza, which is known to transmit efficiently via respiratory droplets. One day later, uninfected ferrets were housed in cages next to the infected animals. Ferrets infected with either of the influenza viruses showed clinical signs of disease and high virus levels in nasal swabs collected over multiple days. However, only ferrets exposed to the H1N1-infected group showed signs of clinical disease, indicating that the cow influenza virus does not transmit efficiently via respiratory droplets in ferrets.
Typically, avian and human influenza A viruses do not attach to the same receptors on cell surfaces to initiate infection. The researchers found, however, that the bovine HPAI H5N1 viruses can bind to both, raising the possibility that the virus may have the ability to bind to cells in the upper respiratory tract of humans.
“Collectively, our study demonstrates that bovine H5N1 viruses may differ from previously circulating HPAI H5N1 viruses by possessing dual human/avian-type receptor-binding specificity with limited respiratory droplet transmission in ferrets,” the authors said.
The National Institute of Allergy and Infectious Diseases (NIAID), part of the National Institutes of Health, funded the work of the University of Wisconsin-Madison researchers.
A Eisfield et al . Pathogenicity and transmissibility of bovine H5N1 influenza virus in mice and ferrets. Nature DOI: 10.1038/s41586-024-07766-6 (2024).
Lauren Byrd-Leotis, Ph.D., with the Viral Respiratory Diseases Section of NIAID’s Division of Microbiology and Infectious Diseases, is available to discuss the findings.
To schedule interviews, please contact the NIAID News and Science Writing Branch, (301) 402-1663, [email protected] .
NIAID conducts and supports research — at NIH, throughout the United States, and worldwide — to study the causes of infectious and immune-mediated diseases, and to develop better means of preventing, diagnosing and treating these illnesses. News releases, fact sheets and other NIAID-related materials are available on the NIAID website .
About the National Institutes of Health (NIH): NIH, the nation's medical research agency, includes 27 Institutes and Centers and is a component of the U.S. Department of Health and Human Services. NIH is the primary federal agency conducting and supporting basic, clinical, and translational medical research, and is investigating the causes, treatments, and cures for both common and rare diseases. For more information about NIH and its programs, visit www.nih.gov .
NIH…Turning Discovery Into Health ®
Connect with Us
- More Social Media from NIH
Bird Flu (H5N1) Explained: U.S. Human Infections Rise To 9
- Share to Facebook
- Share to Twitter
- Share to Linkedin
Here’s the latest news about a global outbreak of H5N1 bird flu that started in 2020, and recently spread among cattle in U.S. states and marine mammals across the world, which has health officials closely monitoring it and experts concerned the virus could mutate and eventually spread to humans, where it has proven rare but deadly.
A sign warns of a outbreak of bird flu.
July 15 Officials confirmed a fourth bird flu case in a poultry farm worker on the same Colorado farm from last week, and a fifth suspected case is pending confirmation from the Centers for Disease Control and Prevention. This brings the national number to nine since the first human cases was detected in the state in 2022, with eight of the cases reported this year.
July 12 Colorado authorities announced three workers in a commercial egg operation have presumptive positive cases of bird flu, bringing the state’s count of known human infections up to as high as five—and the national number to seven—though authorities said none of the workers have been hospitalized, and showed “common respiratory infection symptoms” and pink eye. It’s the first time multiple cases have been recorded at the same location.
July 3 Colorado health officials confirmed the state’s second human case of bird flu in a dairy farm worker who has since recovered and only had mild symptoms, reporting pink eye.
June 25 Finland said it plans to begin vaccinating vulnerable populations like farm workers against bird flu as early as next week using 10,000 vaccine series—each with two doses—acquired as part of a European Union deal with vaccine maker CSL Seqirus to provide up to 40 million vaccines to 15 countries.
June 11 The World Health Organization announced a four-year-old child in India was infected with H9N2 bird flu—a different flu strain from H5N1—but recovered after suffering from seizures, respiratory distress, fever and abdominal cramps; H9N2 has infected around 100 people globally since 1998, and this is the second human case in India.
June 6 Dozens of cows infected with bird flu have either died or been slaughtered in Colorado, Ohio, Michigan, South Carolina and Texas, which is unusual since—unlike poultry—cows cost more to slaughter and around 90% usually make a full recovery, Reuters reported .
June 5 A new study examining the 2023 bird flu outbreak in South America that killed around 17,400 elephant seal pups and 24,000 sea lions found the disease spread between the animals in several countries, the first known case of transnational virus mammal-to-mammal bird flu transmission.
May 30 Another human case of bird flu has been detected in a dairy farm worker in Michigan—though the cases aren’t connected—and this is the first person in the U.S. to report respiratory symptoms connected to bird flu, though their symptoms are “resolving,” according to the CDC.
May 23 A new study with mice suggests that drinking infected milk can spread the disease—and that a certain type of pasteurization may not always be effective in killing the virus.
May 22 Michigan reported bird flu in a farmworker—the second U.S. human case tied to transmission from dairy cows—though the worker had a mild infection and has since recovered.
May 21 Australia reported its first human case of bird flu after a child became infected in March after traveling to India, though the child has since recovered after suffering from a “severe infection,” according to the Victorian Department of Health.
May 16 The USDA conducted a study, and discovered that after high levels of the virus was injected into beef, no trace was left after the meat was cooked medium to well done, though the virus was found in meat cooked to lower temperatures.
May 14 The CDC released influenza A waste water data for the weeks ending in April 27 and May 4, and found several states like Alaska, California, Florida, Illinois and Kansas had unusually high levels, though the agency isn’t sure if the virus came from humans or animals, and isn’t able to differentiate between influenza A subtypes, meaning the H5N1 virus or other subtypes may have been detected.
May 10 The Food and Drug Administration announced it will commit an additional $8 million to ensure the commercial milk supply is safe, while the Department of Agriculture said it will pay up to $28,000 per farm to help mitigate the spread of the disease, totaling around $98 million in funds.
May 9 Some 70 people in Colorado are being monitored for bird flu due to potential exposure, and will be tested for the virus if they show any symptoms, the Colorado Department of Public Health told Forbes—it was not immediately clear how or when the people were potentially exposed.
May 1 The Department of Agriculture said it tested 30 grocery store ground beef products for bird flu and they all came back negative, reaffirming the meat supply is safe.
May 1 The Food and Drug Administration confirmed dairy products are still safe to consume, announcing it tested grocery store samples of products like infant formula, toddler milk, sour cream and cottage cheese, and no live traces of the bird flu virus were found, although some dead remnants were found in some of the food—though none in the baby products.
April 30 Wenqing Zhang, head of WHO's Global Influenza Programme, said during a news briefing "there is a risk for cows in other countries to be getting infected," with the bird flu virus, since it’s commonly spread through the movement of migratory birds.
April 29 The Department of Agriculture told Forbes it will begin testing ground beef samples from grocery stores in states with cow outbreaks, and test ground beef cooked at different temperatures and infected with the virus to determine if it's safe to eat.
April 24 The USDA said cow-to-cow transmission may be occurring due to the cows coming into contact with raw milk—and warned against humans and other animals, including pets, consuming unpasteurized milk to prevent potential infection.
April 18 Jeremy Farrar, chief scientist for the World Health Organization, said during a press conference the threat of bird flu spreading between humans was a “great concern,” since it’s evolved and has increasingly been infecting mammals (on land and sea), which means it could possibly spread to humans.
April 1 The CDC reported the second U.S. human case of bird flu in a Texas dairy farmer who became infected after contracting the virus from infected dairy cows, but said the person was already recovering.
Get Forbes Breaking News Text Alerts: We’re launching text message alerts so you'll always know the biggest stories shaping the day’s headlines. Sign up here .
Can Bird Flu Spread Between Humans?
Bird flu doesn’t “transmit easily from person-to-person,” according to the World Health Organization. Bird flu rarely affects humans, and most previous cases came from close contact with infected poultry, according to the CDC. Because human-to-human spread of bird flu poses “pandemic potential,” each human case is investigated to rule out this type of infection. Though none have been confirmed, there are a few global cases—none in the U.S.—where human-to-human transmission of bird flu was thought to be “probable,” including in China , Thailand , Indonesia and Pakistan .
Is Bird Flu Fatal To Humans?
It is very deadly. Between January 2003 and March 28, 2024 there have been 888 human cases of bird flu infection in humans, according to a report by the World Health Organization. Of those 888 cases, 463 (52%) died. To date, only two people in the U.S. have contracted H5N1 bird flu, and they both were infected after coming into contact with sick animals. The most recent case was a dairy worker in Texas who became ill in March after interacting with sick dairy cows, though he only experienced pink eye. The first incident happened in 2022 when a person in Colorado contracted the disease from infected poultry, and fully recovered.
Is It Safe To Drink Milk Infected With Bird Flu?
Raw, unpasteurized milk is unsafe to drink, but pasteurized milk is fine, according to the FDA. Bird flu has been detected in both unpasteurized and pasteurized milk, but the FDA recommends manufacturers against making and selling unpasteurized milk since there’s a possibility consuming it may cause bird flu infection. However, the virus remnants in pasteurized milk have been deactivated by the heat during the pasteurization process , so this type of milk is still believed safe to consume.
Is It Safe To Consume Meat Infected With Bird Flu?
The CDC warns against eating raw meat or eggs from animals “confirmed or suspected” of having bird flu because of the possibility of transmission. However, no human has ever been infected with bird flu from eating properly prepared and cooked meat, according to the agency. The possibility of infected meat entering the food supply is “extremely low” due to rigorous inspection, so properly handled and cooked meat is safe to eat, according to the USDA. To know when meat is properly cooked, whole beef cuts must be cooked to an internal temperature of 145 degrees Fahrenheit, ground meat must be 160 degrees and poultry must be cooked to 165 degrees. Rare and medium rare steaks fall below this temperature. Properly cooked eggs with an internal temperature of 165 degrees Fahrenheit kills bacteria and viruses including bird flu, according to the CDC. “It doesn’t matter if they may or may not have [avian] influenza… runny eggs and rare pieces of meat” are never recommended, Francisco Diez-Gonzalez, director and professor for the Center for Food Safety at the University of Georgia, told Forbes. To “play it safe,” consumers should only eat fully cooked eggs and make sure “the yolks are firm with no runny parts,” Daisy May, veterinary surgeon with U.K.-based company Medivet, said .
What Are Bird Flu Symptoms In Humans?
Symptoms of bird flu include a fever, cough, headache, chills, shortness of breath or difficulty breathing, runny nose, congestion, sore throat, nausea or vomiting, diarrhea, pink eye, muscle aches and headache. However, the CDC advises it can’t be diagnosed based on symptoms alone, and laboratory testing is needed. This typically includes swabbing the nose or throat (the upper respiratory tract), or the lower respiratory tract for critically ill patients.
How Is Bird Flu Affecting Egg Prices?
This year’s egg prices have increased as production decreased due to bird flu outbreaks among poultry, according to the USDA. A dozen large, grade A eggs in the U.S. costed around $2.99 in March, up almost a dollar from the fall. However, this price is down from a record $4.82 in January 2023, which was also spiked by bird flu outbreaks . Earlier this month, Cal-Maine Foods—the country’s largest egg producer—temporarily halted egg production after over one million egg-laying hens and chickens were killed after being infected with bird flu.
Why Do Poultry Farmers Kill Chickens With Bird Flu?
Once chickens have been infected with bird flu, farmers quickly kill them to help control the spread of the virus, since bird flu is highly contagious and fatal in poultry. The USDA pays farmers for all birds and eggs that have to be killed because of bird flu, as an incentive to responsibly try and curb the spread of the disease. The USDA has spent over $1 billion in bird flu compensation for farmers since 2022, according to the nonprofit Food & Environment Reporting Network.
Is There A Vaccine For The Bird Flu (h5n1)?
The FDA has approved a few bird flu vaccines for humans. The U.S. has a stockpile of vaccines for H5N1 bird flu, but it wouldn’t be enough to vaccinate all Americans if an outbreak were to happen among humans. If a human outbreak does occur, the government plans to mass produce vaccines, which can take at least six months to make enough for the entire population. CSL Seqirus, the maker of one of the approved vaccines, expects to have 150 million vaccines ready within six months of an announcement of a human bird flu pandemic. Although there are approved vaccines for other variants designed for birds, there are none for the H5N1 variant circulating. However, the USDA began trials on H5N1 animal-specific vaccines in 2023.
Key Background
As of May 30, more than 92 million poultry (primarily chickens) in 48 states have been euthanized because of bird flu since 2022, and 57 dairy cow herds across nine states have tested positive, according to data from the CDC (unlike chickens, cows appear to recover from the virus). The USDA believes wild migratory birds are the original source of the cow outbreaks that recently has experts concerned it may mutate and spread more easily in humans, though the CDC said its risk to the public remains low . Farrar called the cattle infections in the U.S. a “huge concern,” urging public health officials to continue closely monitoring the situation “because it may evolve into transmitting in different ways.” The increased number of mammal bird flu infections since 2022 “could indicate that the virus is looking for new hosts, and of course, moving closer to people,” Andrea Garcia, vice president of science, medicine and public health for the American Medical Association, said . The first report of a walrus dying from bird flu was detected in April on one of Norway’s Arctic Islands, and the first U.S. dolphin infected with bird flu died back in 2022, according to a report published April 18. More than 10 human bird flu cases were reported to the World Health Organization in 2023, and all but one survived. Bird flu has devastated bird populations, and 67 countries reported the deaths of 131 million poultry in 2022 alone. Although bird flu typically infects wild birds and poultry, it’s spread to other animals during the outbreak, and at least 10 countries have reported outbreaks in mammals since 2022. Around 17,400 elephant seal pups died from bird flu in Argentina in 2023, and at least 24,000 sea lions died in South America the same year. Besides cattle, bird flu has been detected in over 200 other mammals—like seals, raccoons and bears—in the U.S. since 2022. Although rare, even domestic pets like dogs and cats are susceptible to the virus, and the FDA warns against giving unpasteurized milk to cats to avoid possible transmission.
On June 5, WHO confirmed the first human death of a strain of bird flu that’s never before been seen in humans and is separate from H5N1. A 59-year-old man in Mexico contracted H5N2, and died on April 24 after being hospitalized and developing a fever, diarrhea, nausea, shortness of breath and general discomfort. Cases of H5N2 have been reported in poultry in Mexico, but the man had no history with poultry or animals, WHO said. It’s unclear how he became infected. He was bedridden for weeks prior to the infection, and suffered from several other health conditions.
Further Reading
Another Bird Flu Variant Reaches Humans: What To Know About H5N2—After First-Ever Confirmed Death
WHO Warns Threat Of Bird Flu Spreading To Humans Is ‘Great Concern’ (Forbes)
One In Five Milk Samples From Across US Had Traces Of Bird Flu Virus, FDA Says (Forbes)
Can Pets Get Bird Flu? Here’s What To Know (Forbes)
Avian H5N1 (Bird) Flu: Why Experts Are Worried—And What You Should Know (Forbes)

- Editorial Standards
- Reprints & Permissions
Join The Conversation
One Community. Many Voices. Create a free account to share your thoughts.
Forbes Community Guidelines
Our community is about connecting people through open and thoughtful conversations. We want our readers to share their views and exchange ideas and facts in a safe space.
In order to do so, please follow the posting rules in our site's Terms of Service. We've summarized some of those key rules below. Simply put, keep it civil.
Your post will be rejected if we notice that it seems to contain:
- False or intentionally out-of-context or misleading information
- Insults, profanity, incoherent, obscene or inflammatory language or threats of any kind
- Attacks on the identity of other commenters or the article's author
- Content that otherwise violates our site's terms.
User accounts will be blocked if we notice or believe that users are engaged in:
- Continuous attempts to re-post comments that have been previously moderated/rejected
- Racist, sexist, homophobic or other discriminatory comments
- Attempts or tactics that put the site security at risk
- Actions that otherwise violate our site's terms.
So, how can you be a power user?
- Stay on topic and share your insights
- Feel free to be clear and thoughtful to get your point across
- ‘Like’ or ‘Dislike’ to show your point of view.
- Protect your community.
- Use the report tool to alert us when someone breaks the rules.
Thanks for reading our community guidelines. Please read the full list of posting rules found in our site's Terms of Service.

JESSICA T. SERVEY, LT COL (SEL), USAF, MC, BRIAN V. REAMY, COL, USAF, MC, AND JOSHUA HODGE, CAPT, USAF, MC
Am Fam Physician. 2007;75(3):373-376
Patient information: See related handout on parvovirus B19 , written by the authors of this article.
Author disclosure: Nothing to disclose.
Although most persons with parvovirus B19 infection are asymptomatic or have mild, nonspecific, cold-like symptoms, several clinical conditions have been linked to the virus. Parvovirus B19 usually infects children and causes the classic “slapped-cheek” rash of erythema infectiosum (fifth disease). The virus is highly infectious and spreads mainly through respiratory droplets. By the time the rash appears, the virus is no longer infectious. The virus also may cause acute or persistent arthropathy and papular, purpuric eruptions on the hands and feet (“gloves and socks” syndrome) in adults. Parvovirus B19 infection can trigger an acute cessation of red blood cell production, causing transient aplastic crisis, chronic red cell aplasia, hydrops fetalis, or congenital anemia. This is even more likely in patients with illnesses that have already shortened the lifespan of erythrocytes (e.g., iron deficiency anemia, human immunodeficiency virus, sickle cell disease, thalassemia, spherocytosis). A clinical diagnosis can be made without laboratory confirmation if erythema infectiosum is present. If laboratory confirmation is needed, serum immunoglobulin M testing is recommended for immunocompetent patients; viral DNA testing is recommended for patients in aplastic crisis and for those who are immunocompromised. Treatment is usually supportive, although some patients may require transfusions or intravenous immune globulin therapy. Most patients recover completely.
Parvovirus B19 infection is common worldwide, and most persons who contract the virus are infected by 15 years of age. Infection is most common in late winter or early spring. 1 The virus is transmitted through exposure to infected respiratory droplets or blood products and vertically from mother to fetus. 1 Exposure to respiratory droplets is the most common means of transmission. The transmission rate is about 50 percent for those living with infected persons and about 20 to 30 percent for susceptible teachers and day care workers who are exposed to infected children. 1 Nosocomial transmission also has been documented. 2 The incubation period of the infection ranges from four to 14 days but can last as long as 21 days. 1
| Patients with persistent parvovirus B19 infection may benefit from intravenous immune globulin therapy. | C | |
| Pregnant women who are diagnosed with parvovirus B19 infection should receive serial ultrasonography (weekly or biweekly) for 10 to 12 weeks. | C | |
| Patients presenting with typical erythema infectiosum (fifth disease) do not need laboratory testing to confirm parvovirus B19 infection. | C | , , |
Parvovirus is a small, single-stranded DNA virus. The lack of a lipid envelope makes it resistant to physical inactivation with heat or detergents. 3 The virus targets rapidly growing erythroid progenitor cells, which are found in human bone marrow, fetal liver, human umbilical cord, and peripheral blood. 4 , 5 To become infective, the parvovirus attaches to a P antigen receptor. Persons with parvovirus B19 infection are no longer contagious when the rash appears because viremia has cleared by this point. Most symptoms occur secondary to immune complex formation.
Clinical Conditions Associated with Parvovirus B19
Most persons with parvovirus B19 infection are asymptomatic or exhibit mild, nonspecific, cold-like symptoms that are never linked to the virus. 6 However, clinical conditions associated with the infection include erythema infectiosum; arthropathy; transient aplastic crisis; chronic red cell aplasia; papular, purpuric eruptions on the hands and feet (“gloves and socks” syndrome); and hydrops fetalis. Conditions postulated to have a link to parvovirus B19 infection include encephalopathy, epilepsy, meningitis, myocarditis, dilated cardiomyopathy, and autoimmune hepatitis. 6
ERYTHEMA INFECTIOSUM (FIFTH DISEASE)
Erythema infectiosum is the most recognizable presentation of parvovirus B19 infection. The disease generally affects children four to 10 years of age, 7 although a less-pronounced rash can occur in adults. Prodromal symptoms are mild and include fever, coryza, headache, and nausea. The first stage of the rash ( Figure 1A 8 ) presents as erythema of the cheeks (“slapped-cheek” rash) with circumoral pallor. After one to four days, the second stage ( Figures 1B and 1C 8 ) appears as a maculopapular rash of the extremities and trunk. Central clearing of the rash is possible, giving it a lacy, reticular pattern. 3 The second-stage rash usually lasts one to six weeks. The third stage may continue for the next one to three weeks. The rash persists but varies with exposure to heat or sunlight, 6 resolving spontaneously with no permanent sequelae.
ARTHROPATHY
Arthropathy may be a complication of erythema infectiosum or a primary presentation of parvovirus B19 infection. Approximately 8 percent of children infected with the virus have arthralgia. However, arthralgia is more common in adolescents and adults with parvovirus B19 infection, affecting up to 60 percent of these persons. Arthropathy affects women twice as often as men. 9
The pattern of arthropathy differs between adults and children. In adults, the pattern is symmetric and polyarticular and usually involves the proximal interphalangeal and meta-carpophalangeal joints. It affects the knees, wrists, and ankles less often. Arthropathy generally resolves within three weeks but can last for months to years, especially in women. In children, the pattern can be symmetric or asymmetric and usually involves the knees (82 percent of patients) and ankles. 9 Some patients may test positive for rheumatoid factor and antinuclear antibodies.
There is a significant overlap in symptoms of parvovirus-related arthropathy and those of other diseases (e.g., rheumatoid arthritis, systemic lupus erythematosus). Patients with parvovirus B19 infections do not have articular erosion. 3 , 10
TRANSIENT APLASTIC CRISIS
Persons with decreased erythrocytes caused by conditions such as iron deficiency anemia, human immunodeficiency virus (HIV), sickle cell disease, spherocytosis, or thalassemia are at risk of transient aplastic crisis if infected with parvovirus B19. The virus causes a cessation of erythrocyte production. This can be life threatening, although most patients make a full recovery within two weeks. Multiple blood transfusions may be necessary initially.
The precipitous drop in hemoglobin also may cause congestive heart failure, a cerebrovascular accident, or acute splenic sequestration. White blood cell and platelet counts also may fall. 11 Patients are highly contagious during aplastic crisis and should be isolated to prevent transmission of the virus. 11
CHRONIC RED CELL APLASIA
Parvovirus B19 infection may persist in immunocompromised persons without antibodies. Rashes and arthropathy do not develop because they occur secondary to antibody complex deposition in the skin and joints. 12 Patients present with fatigue and pallor caused by anemia, which can be severe, prolonged, or recurrent. Reticulocytes may be absent and transfusions may be required. If severe anemia continues, intravenous immune globulin treatment may be necessary. 13 The rash and arthropathy may develop secondary to the infusion of antibodies and the formation of immune complexes. Cessation of immunosuppressant or antiretroviral treatment may ameliorate symptoms in patients with HIV. 12
GLOVES AND SOCKS SYNDROME
Parvovirus B19 has been associated with papular, purpuric gloves and socks syndrome, although a causative relationship has not been proven. The syndrome typically occurs in young adults and presents as symmetric, painful erythema and edema of the feet and hands. The condition gradually progresses to petechiae and purpura and may develop into vesicles and bullae with skin sloughing. 14 A hallmark of the syndrome is a sharp demarcation of the rash at the wrists and ankles, although other areas (e.g., cheeks, elbows, knees, inner thighs, glans penis, buttocks, or vulva) may be involved. 15 Patients may generally appear well but may experience arthralgia, fever, or both. Symptoms usually resolve within one to three weeks without scarring. Gloves and socks syndrome also has been associated with hepatitis B, cytomegalovirus, Epstein-Barr virus, human herpesvirus 6, measles, cox-sackievirus B, and drug reactions. 16
HYDROPS FETALIS
Pregnancy does not alter parvovirus B19 infection in the mother, 17 although the fetal liver and heart may become infected. The infant may develop severe anemia, caused by an already shortened red cell lifespan, or may develop myocarditis from direct infection of the heart. The combination of severe anemia and myocarditis can cause congestive heart failure and hydrops fetalis. 18 The estimated risk of transplacental infection is 30 percent. Many fetuses are born without symptoms, but there is a 2 to 6 percent risk of fetal loss. 1 Second-trimester pregnancies are the most vulnerable because of increased hematopoiesis in the liver. Although the placenta has an abundance of P antigen receptors for the virus, first-trimester pregnancies have the lowest risk because of the fetal inability to produce immunoglobulin M (IgM) and the difficulty of antibody transfer across the placenta. 18
If a pregnant woman is exposed to parvovirus B19, acute infection should be confirmed by testing for the presence of IgM antibodies or by seroconversion of IgG antibodies. 19 If acute infection is confirmed, serial ultrasonography (weekly or biweekly) should be performed for 10 to 12 weeks after initial infection to prevent hydrops fetalis. 19 The risk virtually disappears after 12 weeks. If hydrops occurs, fetal blood sampling and possible transfusion are necessary. 19 Routine testing for parvovirus is not indicated in pregnant women. 20
If erythema infectiosum is present, a clinical diagnosis can be made without laboratory testing. 1 , 3 , 7 If laboratory testing is needed, there are two types of diagnostic tests to confirm parvovirus B19 infection: B19-specific antibody testing and viral DNA testing. Giant pronormoblasts on a peripheral blood smear or in a bone marrow aspirate are suggestive of parvovirus B19 infection but are not diagnostic. 21
Serum IgM testing is recommended to diagnose acute viral infection in immunocompetent patients, with 89 percent sensitivity and 99 percent specificity. 22 Elevated IgM antibodies will remain detectable for two to three months after acute infection. IgG testing is less useful because it only indicates previous infection and immunity. 1 Viral DNA testing is crucial for the diagnosis of parvovirus B19 infection in patients in transient aplastic crisis or in immunocompromised patients with chronic infection. These patients do not test positive for IgM or IgG and remain contagious. 11 Polymerase chain reaction (PCR) assays are preferred over less sensitive nucleic acid hybridization assays. The sensitivity and specificity of PCR assays vary widely among laboratories, and, overall, PCR does not appear to be more sensitive than IgM antibody assays for the diagnosis of acute parvovirus infection.
Generally, erythema infectiosum is self-limited and does not require treatment. Patients with arthralgia may require nonsteroidal anti-inflammatory drug treatment. 6 Patients in transient aplastic crisis may require erythrocyte transfusions while the marrow recovers. 12 Chronic red cell aplasia, if severe, may require intravenous immune globulin therapy. 13 This treatment may improve anemia symptoms, but it may precipitate a rash or arthropathy. Intravenous immune globulin also has been used in several case reports of severe illness. 13 A vaccine has been developed but is not yet available. 23
Parvovirus B19 (erythema infectiosum, fifth disease). In: Red Book 2006: Report of the Committee on Infectious Diseases. 27th ed. Washington, D.C.: American Academy of Pediatrics, 2006:484–7.
Bell LM, Naides SJ, Stoffman P, Hodinka RL, Plotkin SA. Human parvovirus B19 infection among hospital staff members after contact with infected patients. N Engl J Med. 1989;321:485-91.
Young NS, Brown KE. Parvovirus B19. N Engl J Med. 2004;350:586-97.
Brown KE, Mori J, Cohen BJ, Field AM. In vitro propagation of parvovirus B19 in primary foetal liver culture. J Gen Virol. 1991;72(pt 3):741-5.
Srivastava CH, Zhou S, Munshi NC, Srivastava A. Parvovirus B19 replication in human umbilical cord blood cells. Virology. 1992;189:456-61.
Heegaard ED, Brown KE. Human parvovirus B19. Clin Microbiol Rev. 2002;15:485-505.
Plummer FA, Hammond GW, Forward K, Sekla L, Thompson LM, Jones SE, et al. An erythema infectiosum–like illness caused by human parvovirus infection. N Engl J Med. 1985;313:74-9.
American Academy of Pediatrics. Red Book Online. Accessed at: http://www.aapredbook.org .
Nesher G, Moore TL. Human parvovirus infection. Infect Med. 1997;14:638-42.
Naides SJ, Scharosch LL, Foto F, Howard EJ. Rheumatologic manifestations of human parvovirus B19 infection in adults. Arthritis Rheum. 1990;33:1297-309.
Smith-Whitley K, Zhao H, Hodinka RL, Kwiatkowski J, Cecil R, Cecil T, et al. Epidemiology of human parvovirus B19 in children with sickle cell disease. Blood. 2004;103:422-7.
Posfay-Barbe KM, Michaels MG. Parvovirus B19 in organ transplant recipients. Curr Opin Organ Transpl. 2003;8:283-7.
Kurtzman G, Frickhofen N, Kimball J, Jenkins DW, Nienhuis AW, Young NS. Pure red-cell aplasia of 10 years' duration due to persistent parvovirus B19 infection and its cure with immunoglobulin therapy. N Engl J Med. 1989;321:519-23.
Alfadley A, Aljubran A, Hainau B, Alhokail A. Papularpurpuric “gloves and socks” syndrome in a mother and daughter. J Am Acad Dermatol. 2003;48:941-4.
Metry D, Katta R. New and emerging pediatric infections. Dermatol Clin. 2003;21:269-76.
Katta R. Parvovirus B19: a review. Dermatol Clin. 2002;20:333-42.
Alger LS. Toxoplasmosis and parvovirus B19. Infect Dis Clin North Am. 1997;11:55-75.
Morey AL, Keeling JW, Porter HJ, Fleming KA. Clinical and histopathological features of parvovirus B19 infection in the human fetus. Br J Obstet Gynaecol. 1992;99:566-74.
American College of Obstetrics and Gynecologists. ACOG practice bulletin. Perinatal viral and parasitic infections. Number 20, September 2000. Int J Gynaecol Obstet. 2002;76:95-107.
Institute for Clinical Systems Improvement. Prenatal care, routine. Accessed June 9, 2006, at: http://www.icsi.org/knowledge/detail.asp?catID=29&itemID=191 .
Cohen BJ, Buckley MM. The prevalence of antibody to human parvovirus B19 in England and Wales. J Med Microbiol. 1988;25:151-3.
Doyle S, Kerr S, O'Keeffe G, O'Carroll D, Daly P, Kilty C. Detection of parvovirus B19 IgM by antibody capture enzyme immunoassay: receiver operating characteristic analysis. J Virol Methods. 2000;90:143-52.
Ballou WR, Reed JL, Noble W, Young NS, Koenig S. Safety and immunogenicity of a recombinant parvovirus B19 vaccine formulated with MF59C.1. J Infect Dis. 2003;187:675-8.
Continue Reading
More in afp, more in pubmed.
Copyright © 2007 by the American Academy of Family Physicians.
This content is owned by the AAFP. A person viewing it online may make one printout of the material and may use that printout only for his or her personal, non-commercial reference. This material may not otherwise be downloaded, copied, printed, stored, transmitted or reproduced in any medium, whether now known or later invented, except as authorized in writing by the AAFP. See permissions for copyright questions and/or permission requests.
Copyright © 2024 American Academy of Family Physicians. All Rights Reserved.
- Open access
- Published: 15 July 2024
Retrospective analysis of hospital electronic health records reveals unseen cases of acute hepatitis with unknown aetiology in adults in Oxfordshire
- Cedric C. S. Tan 1 , 2 ,
- Gavin Kelly 3 ,
- Jack Cregan 4 ,
- Joseph D. Wilson 4 , 5 ,
- Tim James 6 ,
- Meera Chand 7 ,
- Susan Hopkins 8 ,
- Maaike Swets 9 ,
- J. Kenneth Baillie 10 ,
- Katie Jeffery 11 , 12 ,
- Ann Sarah Walker 13 ,
- David W. Eyre 11 , 14 ,
- Nicole Stoesser 4 , 11 &
- Philippa C. Matthews 1 , 4 , 15 , 16
BMC Public Health volume 24 , Article number: 1890 ( 2024 ) Cite this article
28 Accesses
Metrics details
An outbreak of acute severe hepatitis of unknown aetiology (AS-Hep-UA) in children during 2022 was subsequently linked to infections with adenovirus-associated virus 2 and other ‘helper viruses’, including human adenovirus. It is possible that evidence of such an outbreak could be identified at a population level based on routine data captured by electronic health records (EHR).
We used anonymised EHR to collate retrospective data for all emergency presentations to Oxford University Hospitals NHS Foundation Trust in the UK, between 2016–2022, for all ages from 18 months and older. We investigated clinical characteristics and temporal distribution of presentations of acute hepatitis and of adenovirus infections based on laboratory data and clinical coding. We relaxed the stringent case definition adopted during the AS-Hep-UA to identify all cases of acute hepatitis with unknown aetiology (termed AHUA). We compared events within the outbreak period (defined as 1st Oct 2021—31 Aug 2022) to the rest of our study period.
Over the study period, there were 903,433 acute presentations overall, of which 391 (0.04%) were classified as AHUA. AHUA episodes had significantly higher critical care admission rates ( p < 0.0001, OR = 41.7, 95% CI:26.3–65.0) and longer inpatient admissions ( p < 0.0001) compared with the rest of the patient population. During the outbreak period, significantly more adults (≥ 16 years) were diagnosed with AHUA ( p < 0.0001, OR = 3.01, 95% CI: 2.20–4.12), and there were significantly more human adenovirus (HadV) infections in children ( p < 0.001, OR = 1.78, 95% CI:1.27–2.47). There were also more HAdV tests performed during the outbreak ( p < 0.0001, OR = 1.27, 95% CI:1.17–1.37). Among 3,707 individuals who were tested for HAdV, 179 (4.8%) were positive. However, there was no evidence of more acute hepatitis or increased severity of illness in HadV-positive compared to negative cases.
Conclusions
Our results highlight an increase in AHUA in adults coinciding with the period of the outbreak in children, but not linked to documented HAdV infection. Tracking changes in routinely collected clinical data through EHR could be used to support outbreak surveillance.
Peer Review reports
Introduction
In April 2022, the United Kingdom Health Security Agency (UKHSA) alerted the World Health Organization to a significant increase in acute severe hepatitis in children aged less than 10 years, who were otherwise clinically fit and well [ 1 ]. Concerningly, a proportion of these children had sufficiently severe disease to warrant liver transplantation [ 2 ]. Initial investigations and evaluation demonstrated no link to Hepatitis viruses A-E, other known causes of acute hepatitis, toxins, common exposures, or foreign travel; these cases were therefore designated ‘acute severe hepatitis of unknown aetiology’ (AS-Hep-UA).
Subsequent detailed investigation of samples from affected children suggested a likely infectious aetiology, with metagenomic sequencing identifying adeno-associated virus 2 (AAV2) in 81–96% AS-Hep-UA patients (versus 4–7% in controls), alongside a higher than expected prevalence of human adenovirus (HAdV) [ 3 , 4 , 5 ]. In addition to HAdV, a likely contribution was made by AAV coinfection with other ‘helper’ viruses including acute infections or reactivation of latent infections, particularly with Epstein-Barr Virus (EBV), human herpes-virus 6 (HHV6) and enteroviruses [ 3 , 4 , 5 ], and/or a contribution from superantigen-mediated immune activation [ 6 ]. A significant enrichment of the Human Leucocyte Antigen (HLA) class II allele DRB1*04:01 has been identified among AS-Hep-UA cases compared to the background population, suggesting a specific immune susceptibility [ 3 ].
Following the initial reporting of AS-Hep-UA in Scotland, several cases were retrospectively identified in the United States dating back to October 2021 [ 6 ]. By the start of July 2022, > 1000 probable cases had been identified worldwide [ 7 ]. The outbreak in Europe peaked between the end of March and early May 2022 (week 12 to 18), and subsequently declined between May and August [ 6 ]. The case definition of AS-Hep-UA was refined to include age < 16 presenting no earlier than October 1st 2021 with an acute hepatitis and deranged serum liver enzymes (alanine transaminase (ALT) or aspartate transaminase (AST) > 500 IU/L) which could not be accounted for by other causes [ 7 ].
Despite AS-Hep-UA being identified worldwide, there were geographical disparities in the incidence of cases and liver transplantation; rates in the UK and across parts of Europe clearly exceeded expected averages, in contrast to no significant deviation from baseline across the US, Brazil, India, and Japan [ 8 , 9 ]. More than a quarter of global cases were identified in the UK, which had a 100-fold relative incidence rate compared to France, despite the countries being geographical neighbours of almost identical population sizes [ 10 ]. However, the relative contribution of enhanced surveillance, population susceptibility, and circulation of any causative agent to these differing rates has remained unclear. Furthermore, patients with acute hepatitis with unknown aetiology, but not meeting the stringent case definitions would not have been reported as AS-Hep-UA cases (i.e. those with ALT and/or AST elevated but both < 500 IU/L; age ≥ 16 years). Therefore, it is not known whether the AS-Hep-UA outbreak was the ‘tip of an iceberg’ of milder cases of disease in the population, and/or cases among older adolescents and adults.
Routinely collected clinical data (e.g. patient diagnoses, liver enzyme and microbiology test results) in the form of electronic health records (EHR) present an opportunity to investigate population trends that could be associated with this outbreak. There is potential to use routine clinical laboratory parameters as a surveillance tool at a population level, for example as a sentinel marker for circulation of an infectious trigger. In this study, we used hospital EHR data from Oxfordshire, UK, to explore trends before, during and after the period of the AS-Hep-UA outbreak. We addressed the following specific aims: (i) to explore any changes in liver enzyme levels in adults and children/adolescents presenting to hospital, and (ii) to determine any changes in incidence and severity of acute hepatitis and HAdV infection.
Data source
We analysed EHR representing children, adolescents and adults presenting as an emergency to Oxford University Hospitals (OUH) NHS Foundation Trust, a large tertiary referral hospital in the South East of England, serving a population of ~ 725,000. Data were accessed through the Infections in Oxfordshire Research Database (IORD) [ 11 ], and were held, accessed and analysed in accordance with NHS standards for data management and protection (more details in Supplementary methods).
In this retrospective cohort study, we reviewed data from 1st March 2016 to 31st December 2022 for all individuals aged 18 months and older presenting to the Emergency Department or acute medical/surgical assessment units at OUH. We recorded subsequent admission to hospital, admission to the Intensive Care Unit (ICU), duration of hospital admission, and mortality during the admission. Data from all individuals meeting these criteria were included in the analysis (we did not apply any exclusion criteria).
Laboratory data
Laboratory data were generated by externally ISO accredited clinical biochemistry and microbiology laboratories at OUH. A full list of laboratory assays and platforms is provided in Suppl methods, and reference intervals for liver enzymes and inflammatory markers are provided in Table 1 . Laboratory data were based on those routinely collected, where a request for ‘liver function tests’ (LFTs) prompts a clinical biochemistry profile comprising alanine transferase (ALT), alkaline phosphatase (ALP), bilirubin and albumin. Additional laboratory investigations were requested at the discretion of the clinical team. Abnormalities in these biomarkers were classified based on the upper limit of normal (ULN) for all ages and both sexes – mild, moderate and severe derangement was defined as up to 2x, 2–5 × and > 5 × ULN, respectively, with the exception of albumin, which was classified as deranged if levels were less than the lower limit of normal (LLN) of 32 g/L.
HAdV testing was undertaken using a PCR-based multiplex test on respiratory samples or using an HadV-specific PCR on whole blood based on specific clinician request, which usually focuses on patients requiring critical care or in immunocompromised patients under the care of haematology/oncology teams.
Classification and definitions
Patients were stratified into three categories based on their ages at presentation: younger children (< 7 years), older children (7–15 years) and adults (≥ 16 years). Epochs were considered as pre-COVID-19 (1st March 2016—10 March 2020), COVID-19 pandemic period (11th March 2020—31st December 2022), and nested within the COVID-19 pandemic period, the AS-Hep-UA outbreak (1st Oct 2021—31 Aug 2022).
We applied the established strict case definition for AS-Hep-UA, as someone < 16 years of age presenting no earlier than 1st October 2021 with an acute hepatitis (ALT and/or AST > 500 IU/L), which cannot be accounted for by other causes [ 7 ]. We additionally applied a more relaxed definition of acute hepatitis of unknown aetiology (AHUA), to identify cases in adults, and also milder cases that would not meet criteria for AS-Hep-UA. We defined AHUA as patients assigned either a primary or secondary diagnostic code from the International Classification of Diseases 10th Revision consistent with hepatitis of an uncertain cause (Table 2 ; Supplementary Methods) or patients with ALT > 2 × ULN. Diagnostic codes were assigned by hospital admission coders following patient discharge, based on national clinical coding standards. We also considered presentations of diagnosed acute or chronic viral hepatitis A-E virus infection as a baseline control, and to ensure these cases were excluded from the AHUA category. We could not apply specific WHO criteria for acute liver failure, as this would require data regarding the duration of the liver injury (requiring liver function tests prior to the admission, and/or follow-up over time which is outside the scope of this analysis).
We recorded cases of viral Hepatitis A-E as controls, as these infections are likely to be screened, diagnosed and recorded in clinical coding in individuals presenting with a clinical/laboratory picture of hepatitis, allowing us to assimilate baseline comparator data for infectious causes of elevated liver enzymes.
Data sharing
Anonymised LFT data were shared with the Summary Analysis of Laboratory Tests (‘SALT’) project [ 12 ], co-ordinated by the UKHSA as part of the UK-wide public health response, to contribute to a national picture of changes in the incidence of deranged liver function (for epidemiology and ongoing surveillance). The complete datasets analysed during the current study are not publicly available as they contain personal data but are available from IORD, subject to the ethical and governance requirements of the database (details in Suppl methods). To protect anonymity, we avoided disaggregation into any category containing < 5 individuals.
Data analysis and statistical testing
Each presentation episode was considered independently; thus individuals may have featured more than once across the study duration. We used the first set of blood tests taken on presentation for analysis. An infecting pathogen was reported if at least one microbiology test was positive. Data were analysed using R v4.1.2 and visualised using ggplot v3.4.0. The code used for all analyses is hosted on GitHub ( https://github.com/cednotsed/iORD_hepatitis.git ). We tested for the presence of a non-monotonic trend using the non-parametric WAVK test [ 13 ], using its implementation in the R package funtimes [ 14 ]. Fisher’s exact tests and Mann–Whitney U tests were performed using the fisher.test and wilcox.test functions in R . Odds ratios (OR) were calculated using conditional maximum likelihood estimation as part of the fisher.test function. An interupted time series analysis was performed to assess changes in the incidence of AHUA or viral hepatitis A-E during the study duration, using a segmented regression framework [ 15 ], as follows:
Where y t , β i , α outbreak , t start and ε t represent the incidence at time t, the parameter estimates, a binary variable encoding the AS-Hep-UA epoch, the start of the AS-Hep-UA epoch, and model residuals respectively. Autocorrelation and normality of ε t was assessed using the Durbin Watson test in the lmtest R package [ 16 ], and Shapiro–Wilk test, respectively. The statistical significance of parameter estimates was assessed using a Student’s t-test.
Associations between HAdV infection and routinely collected blood biomarker data were assessed using Fisher’s exact test. In particular, for each we tested if the proportion of patients falling into each derangement category (described above) differed significantly between patients with HAdV infections or otherwise. Benjamini–Hochberg procedure was used to correct for multiple testing and adjusted p-values, where available, were annotated.
No evidence for an increase in hospital presentations or elevated liver enzymes during AS-Hep-UA outbreak
We analysed data for 903,433 acute hospital presentations, from 441,780 males and 461,632 females (and 21 individuals for whom sex was not recorded). The median age was 44 years (IQR 22–69 years), with 7.9%, 9.5%, and 82.6% classified as younger children, older children/adolescents, and adults, respectively (Table 1 ). A median of 11,023 patients presented to the hospital per month, with a marked decline in the number of presentations in April 2020 coinciding with the implementation of SARS-CoV-2 (COVID-19) pandemic lockdown measures in the UK introduced on 26th March 2020 (Supplementary Fig. 1a).
During the AS-Hep-UA outbreak, minimal changes in the number of acute presentations per month was observed across any of the three age groups (all WAVK tests p > 0.05; Supplementary Fig. 1a). There was an overall increasing trend in the number of ALT tests requested for acutely presenting patients over time since March 2016 regardless of sex ( WAVK statistic = 13.436, p < 0.0001; Supplementary Fig. 1b). However, there was also an increasing trend in the number of ALT observations compared to WBC observations, which indicates increased ‘liver function scrutiny’ over time (Supplementary Fig. 1c; WAVK statistic = 91.127, p < 0.0001).
Across the study duration, 59% of patient episodes had recorded blood tests. Among these, 90% had an ALT test and 1.2% an AST test. There was no evidence of change in the median or IQR of ALT levels over time for any age group (Fig. 1 a; WAVK tests p > 0.05), with no observable peak during the AS-Hep-UA outbreak. Similarly, the proportion of individuals with mild, moderate or severe derangement of ALT levels remained relatively stable over time (Fig. 1 b; WAVK tests p > 0.05). Therefore, there was no temporal association between the period of the AS-Hep-UA outbreak and elevated liver enzymes in patients presenting acutely to hospital.

Temporal trends in ALT levels in patients presenting acutely to OUH ( a ) median, 25th and 75. th percentiles of ALT levels in patients presenting to OUH and ( b ) proportion of patients with mild, moderate and severe derangement in ALT levels aggregated at 2 month intervals. Relevant epochs are highlighted in grey (pre-COVID-19-pandemic), yellow (COVID-19 pandemic), and with dashed lines (start of AS-Hep-UA outbreak to end of first quarter of 2022)
Increased incidence of AHUA in adults coinciding with AS-Hep-UA outbreak
We further investigated the use of primary or secondary diagnostic codes for identifying increased incidence of AHUA in the patient population. We compared the temporal trend for AHUA diagnoses (Table 2 ; Supplementary Methods) against those for viral hepatitis A-E, the latter which are likely to be relatively stable and therefore serve as an appropriate control.
Across the study duration, 3729 diagnostic codes representing AHUA (total 1005) or viral hepatitis A-E (total 2724) were assigned to 1531 distinct patient episodes (Table 2 ), of which 98% were adults (eight younger children, 11 older children/adolescents, 1512 adults). Overall, there were 391 patient episodes where only diagnostic codes classified as AHUA were assigned, representing 0.04% of all 903,433 patient episodes. The number of acute hepatitis diagnoses classified as AHUA or viral hepatitis A-E per month remained relatively constant over time (Fig. 2 a). However, an increase in the number of AHUA cases coinciding with the AS-Hep-UA outbreak period was observed (Fig. 2 b). The proportion of patients diagnosed with AHUA was higher during the AS-Hep-UA outbreak than outside this period (Fisher’s exact test p < 0.0001; OR 3.01, 95% CI:2.20–4.12).

Temporal trends of acute hepatitis with unknown aetiology (AHUA) based on clinical coding at Oxford University Hospitals from 2016 to 2022. Twelve-month moving averages (means) of ( a ) overall number of hepatitis-related diagnoses (viral hepatitis A-E or AHUA) per month regardless of age group or sex, and ( b ) liver-related diagnoses with or without a specified causal agent (AHUA). ICD10 codes (primary or secondary) and their described causal agents are annotated. Codes are also expanded in Table 2 . Relevant epochs are highlighted in grey (pre-COVID-19-pandemic), yellow (COVID-19 pandemic), and with dashed lines (start of AS-Hep-UA outbreak to end of first quarter of 2022)
Interrupted time series analysis indicated a significantly increased incidence of AHUA (estimate = 2.92, 95% CI: 1.57–5.69; t = 3.50, d.f. = 78, p < 0.001), but not viral hepatitis A-E (estimate = -0.785, 95% CI: -4.34–2.77; t = -0.440, d.f. = 78, p > 0.05), during the AS-Hep-UA outbreak period. Overall, these observations suggest an increased incidence of AHUA amongst adults during the AS-Hep-UA outbreak. We could not determine if this was the case for children, since only 2% of relevant diagnostic codes were assigned to children.
AHUA associated with critical care admission, duration of hospitalisation and mortality
Compared to patients without AHUA, patients with AHUA had significantly higher ICU admission rates (Table 3 ; Fisher’s exact test p < 0.0001; OR 41.7 within and 23.7 outside the AS-Hep-UA epoch) and mortality rates (Fisher’s exact tests p = 0.035 within and p < 0.0001 outside; OR 6.99 within and 11.3 outside), and longer hospitalisations (both Mann–Whitney U tests p < 0.0001; Table 3 ). We also compared patients with AHUA to those diagnosed with viral hepatitis A-E; those with AHUA had significantly higher ICU admission rates (both Fisher’s exact test p < 0.0001; OR 5.01 within AS-Hep-UA outbreak and 3.90 outside outbreak) and longer hospitalisation periods (Mann–Whitney U tests p < 0.05), both within and outside the AS-Hep-UA epoch. Inpatient mortality was significantly higher for the AHUA group than those with viral hepatitis A-E outside the AS-Hep-UA epoch (Fisher’s exact test p < 0.0001; OR 19.8, 95% CI: 4.29–185), but not within the AS-Hep-UA epoch ( p > 0.05; OR = 1.16, 95% CI: 0.0825–16.3).
Increased incidence of HAdV infections during AS-Hep-UA outbreak but not associated with deranged liver enzymes or poorer patient outcomes
Across the study duration, we retrieved 3707 distinct patient records that included microbiology tests for HAdV infection, of which 179 were positive (4.8%). The positivity rate was highest in younger children, among whom 124/781 (15.9%) of HAdV tests were positive, compared to older children/adolescents (9/440, 2.0% positive) and adults (46/2486, 1.9% positive), in keeping with the known epidemiology of HAdV infection [ 17 , 18 , 19 ]. None of the HAdV-infected patients were given ICD10 codes indicative of AHUA across the study duration. A minority (16/179; 9%) of HadV positive results were derived from eye swabs, which is unlikely to have influenced any overall trends.
There was an increase in the number of HAdV-tests undertaken between April 2021 and April 2022 (Fig. 3 a), and a significantly higher number of HAdV tests performed relative to all microbiology tests performed during the AS-Hep-UA epoch (Fisher’s exact test p < 0.0001; OR 1.27, 95% CI:1.17–1.37). These findings indicate increased clinician scrutiny for HAdV during the outbreak. The proportion of HAdV-positive tests during the AS-Hep-UA epoch was significantly higher than outside of the AS-Hep-UA epoch at 60/839 (7.2%) vs 119/2868 (4.1%) respectively (Fisher’s exact test p < 0.001; OR 1.78, 95% CI:1.27–2.47). Additionally, there was an increase in the incidence and proportion of HAdV-positive tests in younger children during the AS-Hep-UA outbreak relative to the period preceding the outbreak (Fig. 3 b). However, there were also other peaks in the proportion of HAdV-positive tests across the entire study duration (Fig. 3 b), indicating previous periods of high HAdV-positivity before the AS-Hep-UA epoch.

Temporal trends of HAdV-related microbiological tests requested at OUH from 2016 to 2022. a Number of HAdV tests requested and b the proportion of all HadV tests that were positive per month. Relevant epochs are highlighted in grey (pre-COVID-19-pandemic), yellow (COVID-19 pandemic), and with dashed lines (start of AS-Hep-UA outbreak to end of first quarter of 2022). Red, green, blue and black lines show data for for younger children (< 7 years), older children/adolescents (7–15 years), adults (≥ 16), and 12-month simple moving average, respectively
There was no evidence that the proportion of patients with mild, moderate or severe derangement of ALT, AST, bilirubin, GGT, CRP or WBC differed significantly between those testing positive vs. negative for HAdV (Fisher’s exact test p > 0.05; Supplementary Fig. 2). The proportion of patients with low albumin and mild derangement of ALP was significantly smaller for those testing HAdV-positive vs. negative (Fisher’s exact test p = 0.01 and p = 0.004 respectively; Supplementary Fig. 2). A similar association with raised ALP was also present in confirmed rhinovirus/enterovirus infections (Fisher’s exact test p = 0.004), indicating that mild derangement of ALP is not unique to HAdV infections (data not shown).
HAdV testing focuses primarily on a clinically vulnerable group, shown by higher rates of hospital admission, ICU admission and inpatient mortality among those receiving a HAdV test (irrespective of the test result) compared to the untested population (78.5% vs 30.7%, 18.2% vs 1.2%, and 0.59% vs 0.34%, respectively; Table 3 ). However, the HAdV-positive group fared somewhat better than those who tested negative, with lower hospital admission (both Fisher’s exact tests p < 0.001) and significantly shorter hospital stays (both Mann–Whitney U tests p < 0.05), whether within or outside the AS-Hep-UA epoch. Additionally, the HAdV-positive group had significantly lower ICU admission rates within the AS-Hep-UA epoch, but not outside the epoch (Fisher’s exact tests p = 0.015 and p = 0.35, respectively) (data not shown). No HAdV-positive patients died across the entire study duration. Characteristics of the population testing positive for HAdV are presented in Table 4 .
We identified an increased incidence of episodes coded as AHUA in adults and an increased incidence of HAdV infections in younger children coinciding with the AS-Hep-UA outbreak in children during 2022. While the latter may be partially accounted for by increased clinician scrutiny during this period, the pattern was not observed to the same level in older children/adolescents, and not at all in adults despite similar increases in testing for all age groups. There was no evidence for increased incidence of abnormal liver enzymes in children or adults, nor associations between HAdV infections and elevated liver transaminases. Our findings suggest that the use of routinely collected liver enzyme EHR data lacks sensitivity for tracking this outbreak, which is likely due a large number of confounding aetiologies that may lead to elevated transaminases. However, the identification of increased incidence of AHUA in adults, which was largely ignored during the AS-Hep-UA outbreak in children, highlights the potential of using hospital diagnostic codes for cost-effective disease surveillance.
Among patients presenting acutely for hospital-based care, HAdV testing is largely reserved for vulnerable groups. Even in this high-risk population, those testing positive for HAdV had lower admission rates than those testing negative, reinforcing the view that this virus is generally benign and self-limiting, with a low risk of serious complications. The lack of associations between HAdV infections and deranged liver enzymes is concordant with the fact that HAdV infections typically lead to mild respiratory or gastrointestinal disease, and that hepatitis is an unusual complication [ 19 ]. Co-infections with HAdV and other viruses such as respiratory syncytial virus (RSV) have been linked to poorer outcomes [ 19 ], but the small number of HAdV infections identified in this study precluded robust analysis of mixed infections. Despite an increased incidence of HAdV infections during the AS-Hep-UA outbreak, this was not an unusual aspect of local HAdV epidemiology, and could have been partly accounted for by increased clinician scrutiny.
It has emerged that severe clinical outcomes during the AS-Hep-UA outbreak in children was likely driven by AAV2 as a leading aetiological agent. However, AAV2 requires co-infection with a ‘helper’ virus (including, but not limited to, HAdV) to replicate, and this appears to be a requirement for the development of liver pathology [ 3 , 4 , 5 ]. Retrospective analysis of EHRs cannot be used to investigate the epidemiology of AAV infection, as these viruses are not part of clinical diagnostic testing pathways and have only been identified as an agent of AS-Hep-UA through retrospective metagenomic sequencing [ 3 , 4 , 5 ]. The specific subtype of HAdV implicated in the outbreak, 41F, is not routinely discriminated from other types by clinical testing. Other possible ‘helper viruses’ include human herpesviruses [ 5 ], many of which are ubiquitous in the population and characterised by long-term carriage and latency, making it difficult to distinguish between clinically relevant episodes and subclinical reactivation. Thus routinely-collected clinical datasets do not include screening for all relevant pathogens, and detection of implicated pathogens can be difficult to interpret due to the detection of commensal or bystander organisms. Overall, wider adoption of metagenomics-based diagnostics has the potential to further enhance the utility of EHRs to investigate future outbreaks but interpretation is complex [ 4 ].
Although we have captured data for a large population over a period of almost seven years, this remains a small data set within which to identify rare events, and our region was not known to be directly affected by the AS-Hep-UA outbreak. Measurement of liver enzymes is a blunt approach to the identification of liver disease, and abnormalities reflect diverse pathology. In the intensive care population, some cases meeting our criteria for AHUA may be related to liver ischaemia or injury from diverse causes associated with critical illness, rather than liver-specific pathology. Hospital coding data varies in its accuracy, and the sensitivity and specificity of codes used to identify cases of hepatitis are not known, but may be low, particularly when the aetiology has not been confirmed.
There are various caveats to our approach for detection of AHUA. If there were milder cases of disease in the population during or preceding the documented AS-Hep-UA outbreak, these may not have presented to hospital at all. Even among those presenting to healthcare, many patients did not have liver enzymes measured. Thus, the IORD dataset provides only a limited view of the whole population, which is not true community surveillance. Hospital admission data are also biased by repeated representation of the same individuals, and over-represent populations who preferentially present to emergency care rather than accessing primary care, or those admitted through different routes. As we did not include children < age 18 months, we may have missed relevant signals in the younger population. Clinical datasets are always subject to missingness, which may not be random. Detection of relevant pathogens also depends on the sample type collected. Our analyses also relies on consistent clinical coding of patient episodes, which is potentially subject to some inaccuracy and/or variability.
Anonymised clinical data from EHRs offers access to large datasets, providing power in numbers to determine overall trends reflecting clinical epidemiology and its influence on morbidity, mortality and health service workload. Monitoring of EHRs may be an effective and low-cost surveillance tool that allows identification of trends that could be of concern—e.g. deranged laboratory parameters and/or changes in recorded diagnoses based on microbiology tests or coding. Such strategies could potentially be developed to provide an ‘early warning’ system to allow clinical and public health authorities to review data in real time, cross-compare between regions, identify possible outbreaks, and implement enhanced surveillance and public-health messaging if necessary. As we have demonstrated, analysis of EHRs allowed us to identify an increased incidence of AHUA amongst adults, which was not determined during the AS-Hep-UA outbreak and warrants further investigation. Longer-term collation of data from multiple regions would offer a more powerful approach that could be extended to other diseases. However, surveillance through EHRs requires the establishment of suitable and systematic data-processing infrastructure and governance frameworks, in addition to investment of personnel and resources, if it is to become a real-world surveillance tool.
Availability of data and materials
The data that support the findings of this study are available from the IORD database but restrictions apply to the availability of these data, which were used under license for the current study, and so are not publicly available. The IORD database can only be accessed by named investigators in accordance with NHS standards for data management and protection. For further details on how to apply for access to the data and for a research proposal template please email [email protected].
All custom scripts used for the analyses presented in this manuscript are hosted on GitHub ( https://github.com/cednotsed/iORD_hepatitis.git ).
World Health Organization, Disease Outbreak News: Acute hepatitis of unknown aetiology – the United Kingdom of Great Britain and Northern Ireland. 2022. https://www.who.int/emergencies/disease-outbreak-news/item/2022-DON368# .
Morfopoulou S, Buddle S, Montaguth OET, Atkinson L, Guerra-Assunção JA, Marjaneh MM, et al. Genomic investigations of unexplained acute hepatitis in children. Nature. 2023. https://doi.org/10.1038/s41586-023-06003-w .
Article PubMed PubMed Central Google Scholar
Ho A, Orton R, Tayler R, Asamaphan P, Herder V, Davis C, et al. Adeno-associated virus 2 infection in children with non-A-E hepatitis. Nature. 2023. https://doi.org/10.1038/s41586-023-05948-2 .
Servellita V, Gonzalez AS, Lamson DM, Foresythe A, Huh HJ, Bazinet AL, et al. Adeno-associated virus type 2 in US children with acute severe hepatitis. Nature. 2023. https://doi.org/10.1038/s41586-023-05949-1 .
Matthews PC, Campbell C, Săndulescu O, Matičič M, Ruta SM, Rivero-Juárez A, et al. Acute severe hepatitis outbreak in children: A perfect storm. What do we know, and what questions remain? 2022.
Kambhampati AK. Trends in acute hepatitis of unspecified etiology and adenovirus stool testing results in children—United States, 2017–2022. MMWR Morb Mortal Wkly Rep. 2022;71(24):797–802.
Article CAS PubMed Google Scholar
Zhang L-Y, Huang L-S, Yue Y-H, Fawaz R, Lim JK, Fan J-G. Acute hepatitis of unknown origin in children: early observations from the 2022 outbreak. J Clin Transl Hepatol. 2022;10:522–30.
van Beek J, Fraaij PL, Giaquinto C, Shingadia D, Horby P, Indolfi G, et al. Case numbers of acute hepatitis of unknown aetiology among children in 24 countries up to 18 April 2022 compared to the previous 5 years. Eurosurveillance. 2022;27:2200370.
PubMed PubMed Central Google Scholar
European Centre for Disease Prevention and Control report; Increase in severe acute hepatitis cases of unknown aetiology in children. https://www.ecdc.europa.eu/en/increase-severe-acute-hepatitis-cases-unknown-aetiology-children .
Oxford Biomedical Research Centre Infections in Oxford Rsearch Database (IORD). https://oxfordbrc.nihr.ac.uk/research-themes/modernising-medical-microbiology-and-big-infection-diagnostics/iord-about/ .
International Severe Acute Respiratory and emerging Infection Consortium (ISARIC). Summary Analysis of Laboratory Tests (SALT). https://isaric4c.net/salt/ .
Lyubchich V, Gel YR, El-Shaarawi A. On detecting non-monotonic trends in environmental time series: a fusion of local regression and bootstrap. Environmetrics. 2013;24:209–26. https://doi.org/10.1002/env.2212 .
Article Google Scholar
Lyubchich V, Gel YR, Brenning A, Chu C, Huang X, Islambekov U, Niamkova P, Ofori-Boateng D, Schaeffer ED, Vishwakarma S, Wang X. Package ‘funtimes’, Functions for Time Series Analysis. 2023. https://cran.r-project.org/web/packages/funtimes/funtimes.pdf .
Lynch III JP, Kajon AE. Adenovirus: Epidemiology, global spread of novel types, and approach to treatment. vol. 42, Thieme Medical Publishers, Inc.; 2021, p. 800–21.
Cooper R, Hallett R, Tullo A, Klapper P. The epidemiology of adenovirus infections in greater Manchester, UK 1982–96. Epidemiol Infect. 2000;125:333–45.
Article CAS PubMed PubMed Central Google Scholar
Sandkovsky U, Vargas L, Florescu DF. Adenovirus: current epidemiology and emerging approaches to prevention and treatment. Curr Infect Dis Rep. 2014;16:1–8.
Lenaerts L, De Clercq E, Naesens L. Clinical features and treatment of adenovirus infections. Rev Med Virol. 2008;18:357–74.
Probst V, Spieker AJ, Stopczynski T, Stewart LS, Haddadin Z, Selvarangan R, et al. Clinical presentation and severity of adenovirus detection alone vs adenovirus co-detection with other respiratory viruses in US children with acute respiratory illness from 2016 to 2018. J Pediatr Infect Dis Soc. 2022;11:430–9. https://doi.org/10.1093/jpids/piac066 .
Article CAS Google Scholar
Magiorkinis G, Matthews PC, Wallace SE, Jeffery K, Dunbar K, Tedder R, et al. Potential for diagnosis of infectious disease from the 100,000 genomes project metagenomic dataset: recommendations for reporting results. Wellcome Open Res. 2019;4:4.
Download references
Acknowledgements
We would like to acknowledge all parties that have provided data or contributed to the creation and maintenance of the IORD database.
Open Access funding provided by The Francis Crick Institute CCST is supported by doctoral funding from A*STAR. PCM is supported by core funding from the Francis Crick Institute (ref CC2223), a Wellcome fellowship (ref 110110/Z/15/Z), and University College London NIHR Biomedical Research Centre (BRC). PCM receives funding from GSK to support a doctoral student in her team, outside the scope of this paper. ASW is supported by the National Institute for Health Research Health Protection Research Unit (NIHR HPRU) in Healthcare Associated Infections and Antimicrobial Resistance at the University of Oxford in partnership with the UK Health Security Agency (UK HSA) (NIHR200915), by the NIHR Oxford Biomedical Research Centre, has core support from the Medical Research Council UK to the MRC Clinical Trials Unit [MC_UU_12023/22] and is an NIHR Senior Investigator. DWE is supported by a Robertson Fellowship and an NIHR Oxford BRC Senior Fellowship. NS is an Oxford Martin Fellow and holds an NIHR Oxford BRC Senior Fellowship.
Author information
Authors and affiliations.
The Francis Crick Institute, London, UK
Cedric C. S. Tan & Philippa C. Matthews
UCL Genetics Institute, University College London, London, UK
Cedric C. S. Tan
Bioinformatics and Biostatistics, The Francis Crick Institute, London, UK
Gavin Kelly
Nuffield Department of Medicine, University of Oxford, Oxford, UK
Jack Cregan, Joseph D. Wilson, Nicole Stoesser & Philippa C. Matthews
King’s College Hospital NHS Foundation Trust, London, UK
Joseph D. Wilson
Department of Biochemistry, Oxford University Hospitals NHS Foundation Trust, Oxford, UK
United Kingdom Health Security Agency, Colindale, UK
Meera Chand
NIHR Health Protection Research Unit, Imperial College London, London, UK
Susan Hopkins
Leiden University Medical Center, Leiden, Netherlands
Maaike Swets
Roslin Institute, University of Edinburgh, Edinburgh, UK
J. Kenneth Baillie
Department of Infectious Diseases and Microbiology, Oxford University Hospitals, Oxford, UK
Katie Jeffery, David W. Eyre & Nicole Stoesser
Radcliffe Department of Medicine, University of Oxford, Oxford, UK
Katie Jeffery
Oxford University Hospitals NHS Foundation Trust, Oxford, UK
Ann Sarah Walker
Big Data Institute, Nuffield Department of Population Health, University of Oxford, Oxford, UK
David W. Eyre
University College London Hospitals NHS Foundation Trust, London, UK
Philippa C. Matthews
University College London, London, UK
You can also search for this author in PubMed Google Scholar
Contributions
DWE, NS and PCM planned the project and wrote the proposal. CCST performed data analyses and prepared the figures. CCST and PM wrote the primary manuscript text. GK, ASW supported statistical analysis. JC curated and provided interpretation of the IORD database. ASW leads management and governance oversight of IORD. All authors provided intellectual input and contributed to the final version of the manuscript.
Corresponding author
Correspondence to Philippa C. Matthews .
Ethics declarations
Ethics approval and consent to participate.
All data used for this study was pseudonymised prior to its use. The study, as part of the Infections in Oxfordshire Research Database (IORD), was approved by the National Research Ethics Service South Central – Oxford C Research Ethics Committee (19/SC/0403), the Health Research Authority and the national Confidentiality Advisory Group (19/CAG/0144), including provision for use of pseudonymised routinely collected data without individual patient consent. As such, individual patient consent was not obtained for the purposes of this study. Individuals who choose to opt out of their data being used in research are not included in the study. The study sponsor was OUH. All patients were assigned an anonymised ‘cluster ID’, with no identifying details handled by the research team. Only month/year of birth was available rather than specific date of birth. Data were held within a password protected, encrypted database and accessed only by named investigators in accordance with NHS standards for data management and protection. More details can be found at the following URL: https://oxfordbrc.nihr.ac.uk/research-themes-overview/antimicrobial-resistance-and-modernising-microbiology/infections-in-oxfordshire-research-database-iord/ .
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note.
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Supplementary material 1., rights and permissions.
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/ . The Creative Commons Public Domain Dedication waiver ( http://creativecommons.org/publicdomain/zero/1.0/ ) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
Reprints and permissions
About this article
Cite this article.
Tan, C.C.S., Kelly, G., Cregan, J. et al. Retrospective analysis of hospital electronic health records reveals unseen cases of acute hepatitis with unknown aetiology in adults in Oxfordshire. BMC Public Health 24 , 1890 (2024). https://doi.org/10.1186/s12889-024-19292-1
Download citation
Received : 10 November 2023
Accepted : 27 June 2024
Published : 15 July 2024
DOI : https://doi.org/10.1186/s12889-024-19292-1
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
- Acute hepatitis
- Epidemiology
- Electronic health records
- Surveillance
BMC Public Health
ISSN: 1471-2458
- General enquiries: [email protected]
Person in Colorado infected with bubonic plague. Here's what to know.

A person in Pueblo County, Colorado, about 100 miles south of Denver, has been infected with bubonic plague , local health officials said Monday.
Last week, state and local officials identified the case based on preliminary test results . Trysten Garcia, a spokesperson for the Pueblo Department of Public Health and Environment, said in an email Tuesday morning it was bubonic plague. Earlier announcements had been more general, stating it was "plague." The infection was initially reported on Friday. The department said the person was hospitalized for bubonic plague but their condition improved. Because plague is endemic locally, it's hard to pinpoint the source of infection, Garcia said.
Bubonic plague, which wreaked havoc during historic pandemics across Europe and Asia, is caused by the Yersinia pestis bacterium. It is transmitted by fleas often carried by rodents in the wild, including rats and prairie dogs. Plague occurs when infected fleas bite people, or when humans touch or skin an infected animal.
Most human cases in the U.S. have occurred in the Southwest – in northern New Mexico, northern Arizona and southern Colorado – and the West – in California, southern Oregon and far western Nevada. The U.S. has an average of seven human plague cases per year, according to the Centers for Disease Control and Prevention.
In March, a New Mexico man died from plague , state health officials announced. Oregon officials identified a human case of plague in February. The Oregon case likely stemmed from an infected pet cat.
Avian influenza: Fourth bird flu case in US in 2024 appears in Colorado, CDC says
What are the symptoms of bubonic plague?
Symptoms can develop after an incubation period lasting between a day and a week, according to the World Health Organization . Plague typically causes sudden fever and chills, severe headache, muscle aches, nausea and vomiting.
A common symptom is swollen, painful lymph nodes, called buboes which give the most common form of the disease its name, bubonic plague. These manifest in the body as swelling in the armpits, groin and neck, the Mayo Clinic said . Buboes vary in size from less than half an inch to about 4 inches. Bubonic plague doesn’t spread from person to person, the CDC said.
Other forms of plague include pneumonic, which happens when the bacteria infects the lungs; and septicemic, when the bacteria infects the blood. Pneumonic plague can spread when someone breathes air droplets from an infected person. These droplets can remain in the air an hour after an infected person coughs. Pneumonic plague can also occur when the bacteria from untreated bubonic or septicemic plague spreads to the lungs. Septicemic plague doesn’t spread from person to person.
The WHO estimates that 30% to 60% of people die of bubonic plague. Pneumonic plague is fatal when left untreated.
There is no vaccine for plague in the U.S., the CDC said. People who develop symptoms of plague should immediately see a health provider. Antibiotics can treat plague, but a person must be seen immediately to avoid serious illness or death, said Alicia Solis, a communicable disease and emergency preparedness program manager in the Pueblo County public health department, in a statement.
How can you avoid it?
Wild rodents in the American West commonly carry plague. With that in mind, Pueblo County health officials warned people to clear areas where rodents can hide and breed near homes, garages, sheds or recreational areas. People should not let pets hunt or roam in rodent areas, including prairie dog colonies, and they should treat dogs and cats for fleas regularly. Officials warned that flea collars have not been proven effective against plague-infested fleas. Officials also warned people to avoid letting their pets sleep with them.
People should also avoid contact with dead animals. If you must handle sick or dead animals, use insect repellant with 20 to 30% DEET to protect against fleas, officials said. Use a long-handled shovel to pick up an animal and place it in an outdoor garbage bag.
Health news: Ozempic v. Mounjaro? There's a clear winner for weight loss, study finds.
'Black Death'
Plague has been found in all continents except Oceania, but the most endemic countries are the Democratic Republic of Congo, Madagascar and Peru. Historically, the disease decimated global populations. In the 14th century, bubonic plague infamously brought the Black Death pandemic, which caused an estimated 50 million deaths, WHO said. More than a quarter of Europe’s population died, although some estimates are higher. Outbreaks later occurred in China and India.
Past U.S. outbreaks, misperceptions
The U.S. has not seen the same level of deaths from plague as other regions, although outbreaks have occurred over the decades. In 1900, plague was introduced on rat-infested ships that sailed from affected areas, causing epidemics in port cities such as San Francisco, where racist public health policies wrongly blamed the Chinese community for bringing disease and being uniquely susceptible to it, as the journalist David K. Randall wrote in “ Black Death at the Golden Gate. ” In the mid-1920s, Los Angeles had the nation’s largest urban epidemic with more than 30 deaths. In Southern California at the time, public health officials blamed the “Mexican district” just east of downtown.
But plague-infested fleas on urban rat species were to blame, and they’d go on to infect rural rodent species in the West with plague to create a reservoir of infestation that continues to this day.
Colorado confirms four human bird flu infections, fifth suspected
- Medium Text

Sign up here.
Reporting by Devika Nair in Bengaluru and Leah Douglas; Editing by Clarence Fernandez
Our Standards: The Thomson Reuters Trust Principles. , opens new tab

TSMC set to report strong profit; stock pressured by Trump comments
Taiwan Semiconductor Manufacturing Co , the dominant producer of advanced chips used in artificial intelligence (AI) applications, is expected to report a 30% jump in second-quarter profit on Thursday thanks to soaring demand.


Medical Virology Pathogenesis of viral infection
Jan 02, 2020
340 likes | 420 Views
Medical Virology Pathogenesis of viral infection. Dr. Sameer Naji, MB, BCh, PhD (UK) Dean Assistant Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University. Pathogenesis of viral infection. Viral epidemiology.
Share Presentation
- chronic infection
- apparent infection
- persistent infections infections
- slow chronic virus infections

Presentation Transcript
Medical VirologyPathogenesis of viral infection Dr. Sameer Naji, MB, BCh, PhD (UK) Dean Assistant Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University
Pathogenesis of viral infection
Viral epidemiology • Endemic: Disease present at fairly low but constant level • Epidemic: Infection greater than usually found in a population • Pandemic: Infections that are spread worldwide • Infectivity: The frequency with which an infection is transmitted when contact between a virus and host occurs • Disease index: # persons develop disease/ total infected • Virulence: # fatal cases/ total # of cases • Incidence: # of new cases within a specific period of time % • Prevalence: # of cases of a disease that are present in a particular population at a given time
What does a pathogen have to do? • Infect (infest) a host • Reproduce (replicate) itself • Ensure that its progeny are transmitted to another host
Virus route of entry • Horizontal: (person to person) • Inhalation- via the respiratory tract ex. RSV, MMR, VZV, Rhinovirus • Ingestion- via the gastrointestinal tract ex. Hep A, Rota, Astroviruses, Caliciviruses • Inoculation- through skin abrasions; mucous membranes (e.g. sexual transmission); transfusion; injections (e.g. by doctors or via shared syringes in drug abuse); transplants • Vertical : i.e. from mother to fetus • Transplacentalex. CMV, rubella, HIV • Delivery ex. Hep B, Hep C, HSV, HIV, HPV • Breast feeding ex. CMV, Hep B, HIV • Zoonotic(animal to human) • Animal bite ex. Rabies • Insect bite ex. Dengue, West Nile • Animal excreta ex. Hanta, Arena
Sites of virus entry cilliated epithelium, mucus secretion, lower temperature gastric acid, bile salts
Terminology • Incubation period: Time between exposure and first symptom
Terminology • Communicability: Ability of virus to shed into secretions • Localized infection: infection limited to site of entry • Disseminated infection: spread throughout the body • Primary viremia: site of entry > regional LN > blood • Secondary viremia: site of entry > regional LN > blood > organs (liver, spleen) > blood
Primary Replication • Having gained entry to a potential host, the virus must initiate an infection by entering a susceptible cell. • This frequently determines whether the infection will remain localized at the site of entry or spread to become a systemic infection
Secondary Replication • Occurs in systemic infections when a virus reaches other tissues in which it is capable of replication, e.g. Poliovirus (gut epithelium - neurons in brain & spinal cord) or Lentiviruses (macrophages - CNS + many other tissues). • If a virus can be prevented from reaching tissues where secondary replication can occur, generally no disease results.
Localized Infections: Virus: Primary Replication: Rhinoviruses U.R.T. Rotaviruses Intestinal epithelium Papillomaviruses Epidermis Systemic Infections: Virus: Primary Replication: Secondary Replication: Enteroviruses Intestinal epithelium Lymphoid tissues, C.N.S. Herpesviruses Oropharynx or G.U.tract Lymphoid cells, C.N.S.
Spread Throughout the Host • Apart from direct cell-cell contact, there are 2 main mechanisms for spread throughout the host: • via the bloodstream • via the nervous system
Via the bloodstream • Virus may get into the bloodstream by direct inoculation - e.g. Arthropod vectors, blood transfusion or I.V. drug abuse. The virus may travel free in the plasma (Togaviruses, Enteroviruses), or in association with red cells (Orbiviruses), platelets (HSV), lymphocytes (EBV, CMV) or monocytes (Lentiviruses). Primary viraemia usually proceeds and is necessary for spread to the blood stream, followed by more generalized, higher titre secondary viraemia as the virus reaches other target tissues or replicates directly in blood cells
Via the nervous system • spread to nervous system is preceded by primary viraemia. In some cases, spread occurs directly by contact with neurons at the primary site of infection, in other cases via the bloodstream. Once in peripheral nerves, the virus can spread to the CNS by axonal transport along neurons (classic - HSV). Viruses can cross synaptic junctions since these frequently contain virus receptors, allowing the virus to jump from one cell to another
Virulence and cytopathogenicity • Virulence: the ability of the virus to cause disease in infected cell • Persistent infection • Latent infection, lysogeny • Chronic infection • Permissive cells allow production of virions and/or transformation • Virulent viruses Kill target cell and cause disease (productive response) • Nonpermissive cells permits cell transformation only • Abortive infection no virus replication, early viral proteins cause cell death • Cytopathic effect
Cytopathic effects Virus-induced damage to cells • Changes in size & shape • Cytoplasmic inclusion bodies • Nuclear inclusion bodies • Cells fuse to form multinucleated cells • Cell lysis • Alter DNA • Transform cells into cancerous cells • Virokines and viroreceptors: DNA viruses; cell proliferate and avoid host defenses
Cytopathic changes in cells
Patterns of viral infection • Inapparent infection( Subclinical infection) • Apparent infection: • Acute infection • Persistent Infection Chronic infections Latent Infection Slow chronic virus infections
Patterns of viral infection Acute followed by clearing Hepatitis B & C Chronic Infection Acute followed by persistent infection and virus overproduction Slow chronic infection Prions
Chronic Infection • Virus can be continuously detected ; mild or no clinical symptoms may be evident.
Latent infection The Virus persists in an occult, or cryptic, from most of the time. There will be intermittent flare-ups of clinical disease , Infectious virus can be recovered during flare-ups . Latent virus infections typically persist for the entire life of the host
Slow virus infection • A prolonged incubation period, lasting months or years, during which virus continues to multiply. Clinical symptoms are usually not evident during the long incubation period .
Overall fate of the cell • The cell dies in cytocidal infectionsthis may be acute (when infection is brief and self-limiting) or chronic (drawn out, only a few cells infected while the rest proliferate)-Cytocidal effect • The cell lives in persistent infectionsthis may be productive or nonproductive (refers to whether or not virions are produced) or it may alternate between the two by way of latency and reactivation-Steady state infection
Transformation-Integrated infection (Viruses and Tumors) • RNA tumor viruses usually transform cells to a malignant phenotype by integrating their own genetic material into the cellular genome and may also produce infectious progeny. • Retroviruses: • Acute transforming viruses: v-srconcogene mimic cellular genes (proto-oncogene) • Insertional mutagenesis: inappropriate expression of a proto-oncogene adjacent to integrated viral genome • Transactivating factors: tax gene in HTLV-1; turns on cellular genes causing uncontrolled cellular proliferation
DNA tumor virus infections are often cytocidal; thus transformation is associated with abortive or restrictive infections in which few viral genes are expressed. The persistence of at least part of the viral genome within the cell is required for cell transformation. This is accompanied by the continual expression from a number of viral genes. • P53: regulates the cell cycle; functions as a tumor suppressor that is involved in preventing cancer. HPV • pRb: prevent excessive cell growth by inhibiting cell cycle progression until a cell is ready to divide. HPV • Apoptosis • P53: initiate apoptosis, programmed cell death, if DNA damage proves to be irreparable
Types of Viral infections at the cellular level Type Virus production Fate of cell Abortive - No effect Cytolytic + Death Persistent Productive + Senescence Latent - No effect Transforming DNA viruses - Immortalization RNA viruses + Immortalization
Mechanisms of viral cytopathogenesis
Possible consequences to a cell that is infected by a virus • Lytic infections: Result in the destruction of the host cell; are caused by virulent viruses, which inherently bring about the death of the cells that they infect. • persistent infections:Infections that occur over relatively long periods of time, Where the release of the viral particles may be slow and the host cell may not be lysed. • latent infections:Delay between the infection by the virus and the appearance of symptoms. • Transformation: Some animal viruses have the potential to change a cell from a normal cell into a tumor cell which grows without restraint.
- More by User

Viral Pathogenesis
Viral Pathogenesis. Derek Wong http://virology-online.com. Viral Pathogenesis. Viral pathogenesis is the process by which a viral infection leads to disease. Viral pathogenesis is an abnormal situation of no value to the virus.
1.71k views • 32 slides

Pathogenesis of viral infection. ( Foundation Block , Microbiology : 2017). Dr.Malak M. El-Hazmi. OBJECTIVES. definition and levels of viral pathogenesis. cellular level. host level. The immune response to viral infection. The stages of viral infection.
1.05k views • 29 slides

DIAGNOSIS OF VIRAL INFECTION
DIAGNOSIS OF VIRAL INFECTION. An Overview. Diagnostic Methods in Virology. 1. Direct Examination 2. Indirect Examination (Virus Isolation) 3. Serology. Direct Examination. 1. Antigen Detection immunofluorescence, ELISA etc.
2.88k views • 38 slides

chapter 32 Viral Pathogenesis
chapter 32 Viral Pathogenesis. Samar Abu Ouda 20060282. Pathogenesis & Immunopathogenesis. **The signs and symptoms of most viral diseases undoubtedly are the result of cell killing by virus-induced inhibition macromolecular synthesis e.g. poliovirus, Ebola virus.
532 views • 16 slides

PATHOGENESIS OF VIRAL DISEASES
PATHOGENESIS OF VIRAL DISEASES. General virology (4). Infection and Disease. Disease only results when: A significant proportion of essential cells, which cannot be replaced by regeneration, are destroyed
391 views • 10 slides

768 views • 47 slides

Viral Pathogenesis. Dr. Luka Cicin-Sain Dep. Of Vaccinology, HZI Tel. 0531 6181 4616 [email protected]. Teaching Material: Principles of Virology - Molecular Biology, Pathogenesis, and Control of Animal virus SJ Flint, LW Enquist, VR Racaniello & AM Skalka
1.37k views • 59 slides

Medical Virology
Medical Virology . General Virology . Conception Viruses Virion Size and Shape Structure Replication Viral Variation Classification. Conception.
1.88k views • 131 slides

Viral Pathogenesis. Viral Pathogenesis. Viral pathogenesis is the process by which a viral infection leads to disease. Viral pathogenesis is an abnormal situation of no value to the virus.
1.06k views • 32 slides

Viral Pathogenesis. Prof. Dr. Martin Messerle Hannover Medical School Institute of Virology Tel. 0511-532 4320 [email protected]. Teaching Material: Principles of Virology - Molecular Biology, Pathogenesis, and Control of Animal virus
783 views • 52 slides

Diagnosis of Viral Infection
Diagnosis of Viral Infection. A. Specimen collection and transport. 1. Specimen for viral isolation Specimens for the detection of virus should be collected as early as possible following the onset of disease.
1.02k views • 48 slides

PATHOGENESIS OF BACTERIAL INFECTION
PATHOGENESIS OF BACTERIAL INFECTION. PATHOGENICITY VIRULENCE. The pathogenesis of bacterial infection includes the initiation of the infectious process and the mechanisms leading to the development of signs and symptoms of bacterial disease .
1.56k views • 35 slides

Viral Pathogenesis. Pathogenesis: the process by which one organism causes disease in another Two components of viral disease: Effects of virus replication on the host Effects of host response on virus and the host
1.85k views • 38 slides

Introduction to medical virology “ Viral structure and Classification”
Introduction to medical virology “ Viral structure and Classification”. By: Dr. Malak El-Hazmi Dr. Abdulkarim Alhetheel. OBJECTIVES. General characteristics of viruses. Structure & symmetry of viruses. Classification of viruses. Steps of virus replication .
962 views • 41 slides

Virology – Pathogenesis of viral infections JU- 2 nd Year Medical Students
Virology – Pathogenesis of viral infections JU- 2 nd Year Medical Students. By Dr Hamed AlZoubi – Microbiology and Immunology Department – Mutah University. MBBS (J.U.S.T) MSc, PhD medical microbiology (UK). FRCPath (associate, medical microbiology). [email protected].
506 views • 27 slides

Medical Virology Viral Carditis
Medical Virology Viral Carditis. Dr. Sameer Naji, MB, BCh, PhD (UK) Dean Assistant Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University. Myocarditis – Introduction.
267 views • 24 slides

Introduction to medical virology “ Viral structure and Classification”. Dr. Abdulkarim Alhetheel Assistant Professor in Microbiology Unit College of Medicine & KKUH. OBJECTIVES. General characteristics of viruses. Structure & symmetry of viruses. Classification of viruses.
454 views • 41 slides

449 views • 35 slides

Viral Infection of FGT
Viral Infection of FGT. Dr. Tarek Atia. General considerations: Most of the viruses affecting oral mucosa have the ability to infect the vagina. Routine vaginal smears are usually useful for detecting the typical viral cellular changes. Cellular Changes With Viral Infection.
310 views • 24 slides

Introduction to medical virology “ Viral Pathogenesis”
Introduction to medical virology “ Viral Pathogenesis”. Dr. Abdulkarim Alhetheel Assistant Professor in Microbiology Unit College of Medicine & KKUH. OBJECTIVES. Definition and levels of viral pathogenesis. Types of viral infections at cellular level. Pathogenesis at host level.
328 views • 25 slides
IMAGES
VIDEO
COMMENTS
Symptomatic Presentation. Symptoms can be difficult to differentiate from, and can overlap with, other viral respiratory illnesses such as influenza (flu) and respiratory syncytial virus (RSV). COVID-19 can vary from asymptomatic infection to critical illness; symptoms and severity can change during the illness.
Coronavirus disease 2019 (COVID-19) is an illness caused by the virus SARS-CoV-2. Learn more about the symptoms, risks and ways to protect yourself.
Most commonly, viral infections involve the nose, throat, and upper airways, or systems such as the nervous, gastrointestinal, and reproductive systems. Doctors may base the diagnosis on symptoms, blood tests and cultures, or examination of infected tissues. Antiviral drugs may interfere with the reproduction of viruses or strengthen the immune ...
Download the Mosquito Diseases: West Nile Virus presentation for PowerPoint or Google Slides. Taking care of yourself and of those around you is key! By learning about various illnesses and how they are spread, people can get a better understanding of them and make informed decisions about eating, exercise, and...
An infection is the invasion of the body's tissues by microorganisms such as bacteria and viruses. These disease-causing agents produce reactions in their host as the immune system fights the microorganisms and the toxins they produce. A cold is a type of viral infection.
The clinical presentation of COVID-19 ranges from asymptomatic to critical illness. An infected person can transmit SARS-CoV-2, the virus that causes COVID-19, before the onset of symptoms. Symptoms can change over the course of illness and can progress in severity. Uncommon presentations of COVID-19 can occur, might vary by the age of the ...
Share information about how to treat infectious diseases in this editable template for Google Slides and PowerPoint presentations.
Although there is no shortage of information on this pandemic virus presented in everyday practice, this paper presents a comprehensive review of the latest information on SARS-CoV-2 highlighting the epidemiology, pathogenesis, and clinical aspects of SARS-CoV-2 infection.
By swapping the constituent presentation, we present a strategy for rapidly screening cell types and their respective susceptibility to viral infection without the need for virus particles (virus ...
This presentation aims at targeting a similar virus and its infections known as the RSV virus causing respiratory issues in children. The descriptive layout of this template makes it apt for presenting various medical presentations that are impressive and convince enough.
Study suggests reinfections from the virus that causes COVID-19 likely have similar severity as original infection. NIH-funded analysis of health record data shows severe reinfections often follow severe first infections.
Learn about the epidemiology, clinical presentation, management, and infection control measures of SARS, a severe respiratory illness caused by a novel coronavirus.
A(H5N1) virus infection. 2. Describe risk of human infection with HPAI A(H5N1) viruses, identify higher risk populations, and what to assess in clinical settings. 3. Describe testing, using antivirals, and infection prevention and control recommendations for patients with HPAI A(H5N1) virus infection.
Burton's Microbiology for the Health Sciences Chapter 18. Viral Infections. Chapter 18 Outline. Introduction How do Viruses Cause Disease? Infectious Diseases of the Skin Viral Infections of the Ears Viral Infections of the Eyes Viral Infections of the Respiratory System Slideshow 6145683 by...
Viral Infections: an overview Dr. GerrardUy. Defining a Virus • Viruses consist of a nucleic acid surrounded by one or more proteins • obligate intracellular parasites: they can replicate only within cells • Many human viruses are simply composed of a core and a capsid • Genes: contain either DNA or RNA. Viral Infection • Transmission ...
Even though the host employs a multipronged innate antiviral response to restrict or eliminate the virus infection, viruses have evolved their own effective strategies to antagonize such response and multiply.
Bovine HPAI H5N1 virus thus possesses features that may facilitate infection and transmission in mammals. Highly pathogenic H5N1 avian influenza (HPAI H5N1) viruses occasionally infect, but ...
Implement recommended infection prevention and control practices in the facility. Be aware when respiratory virus levels are increasing in the community and ensure access to respiratory viral testing with rapid results. Guidance and recommendations. Preventing Transmission of Viral Respiratory Pathogens in Healthcare Settings
Recurrent infections are caused by reactivation of the virus, which travels back to the skin or mucous membranes resulting in an active infection. Immune mechanisms suppress the virus with clearing of the lesions in 1-2 weeks, but latency in the ganglia persists.
Wealthy nations are purchasing vaccines against H5N1 influenza and boosting surveillance, but there are concerns that low-income countries will be left behind.
Media Advisory. Monday, July 8, 2024. Features of H5N1 influenza viruses in dairy cows may facilitate infection, transmission in mammals. H5N1 virus did not efficiently transmit via respiratory route to ferrets.
Pathogenesis of viral infection. ( Foundation Block , Microbiology : 2017). Dr.Malak M. El-Hazmi. OBJECTIVES. definition and levels of viral pathogenesis. cellular level. host level. The immune response to viral infection. The stages of viral infection.
Bird flu typically spreads among birds, but there have been recent outbreaks among cattle in the U.S., and one Texas man contracted the virus from sick cows. Here's why so many experts are ...
Although most persons with parvovirus B19 infection are asymptomatic or have mild, nonspecific, cold-like symptoms, several clinical conditions have been linked to the virus. Parvovirus B19 ...
Diagnostic codes were assigned by hospital admission coders following patient discharge, based on national clinical coding standards. We also considered presentations of diagnosed acute or chronic viral hepatitis A-E virus infection as a baseline control, and to ensure these cases were excluded from the AHUA category.
Wild rodents in the American West commonly carry bubonic plague. State officials say to avoid contact with animals and to take care of pets.
Determinants of viral disease: Viral factors AND host factors Nature of disease:- Strain of virus (virulence) - Target tissue: where virus enters the body ability of virus to gain access to target tissue viral tropism permissivity of cells Severity of disease: - virus: ability of infection to kill cells (cytotoxic effects); quantity of virus inoculated; duration of virus infection; other ...
During today's U.C. CDC Clinician Outreach and Communication Activity call, the presentation offered insights regarding the multi-year Highly Pathogenic Avian Influenza (HPAI) (H5N1) virus outbreak in the United States. As of July 16, 2024, the HAPI virus is widespread among wild birds and continues to cause outbreaks in poultry and spillover to mammals, including dairy cattle. To date, three ...
Colorado reported four confirmed infections of H5N1 bird flu virus in poultry workers, and is checking on the status of a fifth suspected case, health authorities said on Sunday.
Medical Virology Pathogenesis of viral infection. Dr. Sameer Naji, MB, BCh, PhD (UK) Dean Assistant Head of Basic Medical Sciences Dept. Faculty of Medicine The Hashemite University. Pathogenesis of viral infection. Viral epidemiology.